


Abstract
Background: Picrasma quassioides (PQ) is a traditional Asian herbal medicine with anti-tumor properties that can inhibit the viability of HepG2 liver cancer cells. H-Ras is often mutated in liver cancer, however, the effect of PQ treatment on H-Ras mutated liver cancer is unclear. This study aimed to investigate the role of PQ on ROS accumulation and mitochondrial dysfunction in H-ras mutated HepG2 (HepG2G12V) cells. Materials and Methods: PQ ethanol extract-induced HepG2G12V apoptosis was analyzed by the MTT assay, fluorescence microscopy, flow cytometry and western blotting. Results: PQ treatment affected cell migration and colony formation in HepG2G12V cells. Cleaved-caspase-3, cleaved-caspase-9 and BCL2 associated agonist of cell death (BAD) expression levels were increased, while the levels of B-cell lymphoma-extra large (Bcl-xL) were decreased with PQ treatment. PQ treatment led to a reduction of H-Ras expression levels in liver cancer cells, thus reducing their abnormal proliferation. Furthermore, it led to increased expression levels of Peroxiredoxin VI, which regulates the redox signal in cells. Conclusion: Taken together these results provide a new functional significance for the role of PQ in treating HepG2G12V liver cancer.
Liver cancer is a malignant tumor with extremely high morbidity and mortality, which may be caused by hepatitis virus infection, smoking, alcoholism, and obesity (1). It has a rapid onset and is frequently detected late; this seriously affectes the clinical treatment of patients with liver cancer (2). RAS is a proto-oncogene that regulates the proliferation and differentiation of cancer cells, and its family members are H-Ras, K-Ras and N-Ras (3). Mutations in RAS gene can be found in a variety of tumor tissues. In response to environmental factors or intracellular signal transduction, Ras proteins get activated and can accelerate the development of cancer. It has been reported that RAS genes have a certain degree of mutations in liver cancer (4), pancreatic cancer (5), colon cancer (6), lung cancer (7), thyroid cancer (8) and other cancers (9, 10). In liver cancer, the RAS gene mostly exists in the form of a mutated H-Ras (11). When RAS gene is mutated, it may lead to abnormal cell proliferation, resistance to apoptotic signals and weakened therapeutic effect of commonly used chemotherapy drugs in clinical practice (12). Therefore, we focused on the development of high-efficiency and low-toxic drugs based on natural Asian herbal medicine.
In recent years, many Asian herbal extracts or derivatives that can be used to treat a variety of diseases have been identified by researchers (13). Specifically, these extracts have been used to treat diabetes, hypertension, cancer and other complex diseases (14-16). There have been reports showing that piceatannol could inhibit the invasive phenotype of MCF10A human breast epithelial cells harboring mutated H-Ras (17), Triptolide and Magnolin could induce apoptosis in K-Ras mutated pancreatic cancers cell (18) and ovarian cancer cell (19).
Picrasma quassioides (P. quassioides or PQ) belonging to the Simaroubaceae family, is a traditional Asian herbal medicine. The main chemical components contained in PQ are quassinoids and alkaloids. In recent years, most studies on PQ were focused on its anti-inflammatory, anti-bacterial and anti-hypertensive properties. The anti-tumor effect of PQ, which is limited to the β-carboline enantiomers extracted from PQ, can cause a reduction in HepG2 liver cancer cell viability (20) while Canthin-6-one alkaloids can induce Bel-7402 liver cancer cell death (21). In addition, PQ methanol and ethanol extracts can induce apoptosis in cervical cancer HEp-2 (22) and HeLa (23) cells and Picrasidine G can inhibit the proliferation of MDA-MB468 breast cancer cells (24).
However, the effect of PQ treatment on H-Ras mutated liver cancer is unclear. In this study, we aimed to investigate the mechanism of PQ treatment in H-Ras mutated liver cancer. To this end, we used PQ ethanol extract to treat H-ras mutated HepG2 cells (HepG2G12V cells). Cell viability, apoptosis and apoptosis-related proteins were evaluated. Furthermore, we analyzed changes in intracellular and mitochondrial levels of the Reactive Oxygen Species (ROS). The mitochondrial-dependent signal changes were also measured. Our results may provide a new understanding of PQ extract function in the treatment of liver cancer, as well as new research directions and ideas for the treatment of H-Ras mutated liver cancer.
Materials and Methods
Extraction and dissolution. Picrasma quassioides (PQ) leaves were chopped. The chopped material (50 g) was submerged in ethyl alcohol (500 ml) at 37°C under shaking at 200 rpm for 5 h. We then used these 500 ml of ethyl alcohol to immerse residues and recover the filtrate. This was repeated three times. The extract solution was filtered and concentrated on a rotary evaporator for 12 h at 55°C. After dissolving in water, the extract was frozen at 80°C, and freeze-dried using a lyophilizer. It was ultimately reconstituted in double distilled water (DDW) and DMSO (1:1) in order to obtain a final stock concentration of 10 mg/ml.
Cell culture. Liver carcinoma cells (HepG2), G12V mutation HepG2 cells (HepG2G12V) were obtained from Disease Models Animal Research Center. HepG2 and HepG2G12V cells were cultured in Dulbecco's modified eagle medium (DMEM, Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS, Hyclone, Logan, UT, USA), penicillin (100 U/ml) and streptomycin (100 mg/ml) (P/S) (Solarbio life sciences, Beijing, PR China), and incubated at 37°C and 5% CO2.
MTT assay. HepG2 and HepG2G12V cells (4×104/ml) were seeded in 96-well plates (NEST Biotechnology, Wuxi, Jiangsu, PR China) treated with various concentrations of PQ for 24 h as well as with 5-Fluorouracil (5-Fu, Solarbio life sciences, Beijing, PR China) that was used as a positive control. After 10 μl of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT, Sigma-Aldrich, St. Louis, MO, USA) was added to each well of 96-well plate and incubated for 4h at 37°C. The supernatant was the discarded and 100 μl of DMSO was added in each well. Finally, the plate was measured under a microplate reader at 490 nm.
Colony formation assays. HepG2G12V cell (1.5×103 cells/well) were cultured in 6-well plates (NEST Biotechnology, Wuxi, Jiangsu, PR China), and treated with various concentrations of PQ for seven days. After that, the cells were fixed with paraformaldehyde for 10 min and washed with 1×PBS. Subsequently, after being fixed with methanol for 20 min, the cells were washed with 1×PBS, and then stained with crystal violet for 30 min, and washed with PBS before imaging.
Cell migration assays. Cell migration was evaluated using the uniform linear scratches. The adherent HepG2G12V cells (seeded in 24-well plates, 2.5×105 cells per well) (NEST Biotechnology, Wuxi, Jiangsu, PR China) were scratched with a small pipette tip. Next, PQ was added to DMEM with 1% of FBS and cultured for 24 h. Each experiment group was imaged at 0 h and 24 h.
Reactive oxygen species detection. Cells were seeded into 24-well plates (NEST Biotechnology, Wuxi, Jiangsu, PR China) at 4×104 cell/well and treated with PQ for 24 h. In order to test the ROS production in cells, we used Dihydroethidium (DHE, Beyotime Biotechnology, Shanghai, PR China) staining. Because the mitochondria are the main _target organs of ROS production, the ROS in mitochondria was measured with MitoSOX (Thermo Fisher Scientific, Waltham, MA, USA) staining. Nuclei were visualized using DAPI (Bioss biology, Beijing, PR China) staining. Nuclear staining was observed qualitatively under a fluorescence microscope (EVOS®x) after a 20-min incubation with DAPI.
Mitochondrial membrane potential detection. The mitochondrial membrane potential in cells is measured using the JC-1 fluorescent probe (Beyotime Biotechnology, Shanghai, PR China), a cation-ion lipophilic dye that accumulates in living mitochondria. When the mitochondrial membrane potential (MMP) is high, JC-1 accumulates in the matrix of the mitochondria, forming an aggregate, which can generate red fluorescence. In this way, the change in mitochondrial membrane potential can be detected by the fluorescence transition of aggregates and monomers. Cells were seeded into 24-well plates at 4×104 cell/well and treated with PQ for 24 h. Afterwards, they were incubated with JC-1 for 20 min and were observed under a fluorescence microscope.
Apoptosis detection. Cell apoptosis was analyzed with the Annexin V-APC Apoptosis Detection Kit (Solarbio life sciences, Beijing, PR China). The HepG2G12V cells were stained according to the manufacturer's protocol and analyzed by fluorescence microscopy after PQ treatment.
ATP synthesis Detection. Cells were seeded into 6-well (NEST Biotechnology, Wuxi, Jiangsu, PR China) plates at 2×105 cells/well and treated with PQ for 24 h. Intracellular Adenosine triphosphate (ATP) was determined by the ATP determination Kit (Thermo Fisher Scientific, Waltham, MA, USA). Luminance (RLU) was measured using a fluorescent chemical analyzer (Infinite 200 PRO Microplate Reader, Tecan Trading AG, Männedorf, Switzerland) according to the manufacturer's protocol immediately after adding the reagent.
Western blot analysis. After PQ treatment, cell proteins were extracted with lysis buffer. The protein sample was loaded and separated discontinuously on 12% sodium dodecyl sulfate-polyacrylamide gels (SDS-PAGE) and transferred onto nitrocellulose membranes (Millipore, Bedford, MA, USA). The membranes were blotted with primary antibodies against caspase-3 (Cell Signaling Technology, Beverly, MA, USA, #9661), caspase9 (Cell Signaling Technology, #9505), caspase-8 (Cell Signaling Technology, #9496), Bax (Abcam Cambridge, MA, USA, #ab90435), Bcl-xL (Santa Cruz Biotechnology, Dallas, TX, USA, #sc-8392), H-Ras (Bioss biology, Beijing, PR China, #bs-1071R), Erk (Santa Cruz Biotechnology, #sc-135900), p-Erk (Santa Cruz Biotechnology, #sc-7383), P38 (AbFrontier, Seoul, Republic of Korea, #LF-MA0126), p-P38 (Santa Cruz Biotechnology, #sc-7973), Jnk (Santa Cruz Biotechnology, #sc-7345), p-Jnk (Santa Cruz Biotechnology, #sc-6254), Prx I (Santa Cruz Biotechnology, #sc-7381), Prx II (AbFrontier, #LF-MA0144), Prx III (Santa Cruz Biotechnology, #sc-59661), Prx IV (Santa Cruz Biotechnology, #sc-376668), Prx V (Santa Cruz Biotechnology, #sc-133073), Prx VI (Santa Cruz Biotechnology, #sc-134478) and β-actin (Abcam Cambridge, #ab7291) (dilution, 1:2,000) at 4°C overnight. The membranes were washed five times with tris buffered saline containing Tween-20 (TBST) [10 mM Tris HCl (pH 7.5), 150 mM NaCl and 0.2% Tween-20] and were subsequently incubated with horseradish peroxidase conjugated goat anti-rabbit IgG (Sangon Biotech, Shanghai, PR China) or anti-mouse IgG (Sangon Biotech, Shanghai, PR China) for 1 h at room temperature (RT). After the removal of excess antibodies by washing with TBST, specific binding was detected using a chemiluminescence detection system (GE Healthcare Life Sciences, Chalfont, UK) according to the manufacturer's protocol.
Statistical analysis. Data from at least three independent experiments were described as the means±standard deviation (SD). Repeated measures analysis of variance (ANOVA) was used to analyze changes across time and differences between groups in each experiment. For most experiments, unpaired two-tailed Student t test was used to determine statistical significance among two groups All differences were considered statistically significant if the p-value was less than 0.05 (*p<0.05; **p<0.01; ***p<0.001).
Results
Effect of PQ treatment on HepG2G12V cell apoptosis, migration and colony formation ability. Elevated expression levels of H-Ras have been reported in liver cancer tissues compared with normal tissues (4). Therefore, to compare the expression of H-Ras in liver cancer tissues with normal liver tissues, we used the Gene Expression Profiling Interactive Analysis (GEPIA) website from the The Cancer Genome Atlas (ATGC) database. The results showed that the expression of H-Ras in the liver tissue of liver cancer patients was significantly higher compared to normal liver tissue. At the same time, patients who had low H-Ras expression levels had significantly higher survival rate than patients with high expression of H-Ras (Figure 1A). To investigate the sensitivity of HepG2G12V cells to PQ treatment, we performed in vitro experiments. The cytotoxicity of PQ in various concentrations (0, 20, 40, 60, 80 and 100 μg/ml) and 5-Fu (100 μg/ml) to HepG2G12V cells was detected by MTT Assay. The results showed that with increasing PQ concentration, cell survival rates decreased in a concentration-dependent manner (Figure 1B). We compared the IC50 values of HepG2 and HepG2G12V cells treated by PQ, indicating that the treatment effect of PQ on HepG2G12V cells is not better than HepG2. There was no big difference; this is what we expected to be acceptable. At the same time, we also tested the characteristics of HepG2 and HepG2G12V cells from the detection of H-Ras expression in Western blot analysis to prove our transfection effect on HepG2G12V cells (Figure 1C). In order to detect the mechanism of PQ-induced HepG2G12V cells apoptosis, HepG2G12V cells were treated with PQ (0, 60, 100 100 μg/ml) for 24 h. After that, the cells were stained with Annexin V-APC and detected by fluorescence microscopy. The results showed that the apoptosis in HepG2G12V cells was increased with increasing PQ concentration (Figure. 1D and E). Some characteristics of cancer cells change during apoptosis. In this regard, the migration and colony forming ability of HepG2G12V cells treated with PQ were affected. Cell migration and colony formation ability of HepG2G12V cells decreased with increasing PQ concentration (Figure 1F).
PQ-induced ROS accumulation and mitochondrial dysfunction in HepG2G12V cells. In order to detect the ROS levels of HepG2G12V, cells were treated with PQ (0, 60, 100 μg/ml) for 24 h, labeled with DHE and the fluorescence signal was measured. The results showed that the level of ROS in cells was increased significantly in response to increased PQ concentration (Figure 2A). Mitochondria are the main source of ROS production, and they are also the main _target of ROS-induced cell damage. The production of ROS, which is a by-product of respiration, occurs mainly at the electron transport chain. That leads to excessive accumulation of ROS, which in turn damages mitochondrial membrane potential. We used JC-1 and MitoSOX staining to measure MMP and the level of ROS in cells treated with PQ. The results showed that MMP was decreased in response to increasing PQ concentration (Figure 2B). In the meantime, ROS levels in mitochondria increased with increasing PQ concentration (Figure 2C). Mitochondria provide metabolic energy in the form of ATP through oxidative phosphorylation. When ATP synthesis levels drop, it indicates a damage in mitochondrial function. The measurements of ATP in PQ-treated HepG2G12V cells showed that the level of ATP production in cells treated with PQ decreased (Figure 2D). The damage of mitochondrial function can lead to apoptosis. In order to detect whether PQ can induce mitochondrial-dependent apoptosis in HepG2G12V cells, we tested the apoptosis-related protein changes over different time periods in HepG2G12V cells treated with PQ (100 μg/ml). The results showed that the protein expression levels of cleaved-caspase-3, cleaved-caspase-9 and BAD were increased and Bcl-xL protein levels were decreased in response to PQ treatment in a time-dependent manner. Concurrently, PQ treatment decreased the level of H-Ras expression in HepG2G12V cells in a time-dependent manner (Figure 2E, F).
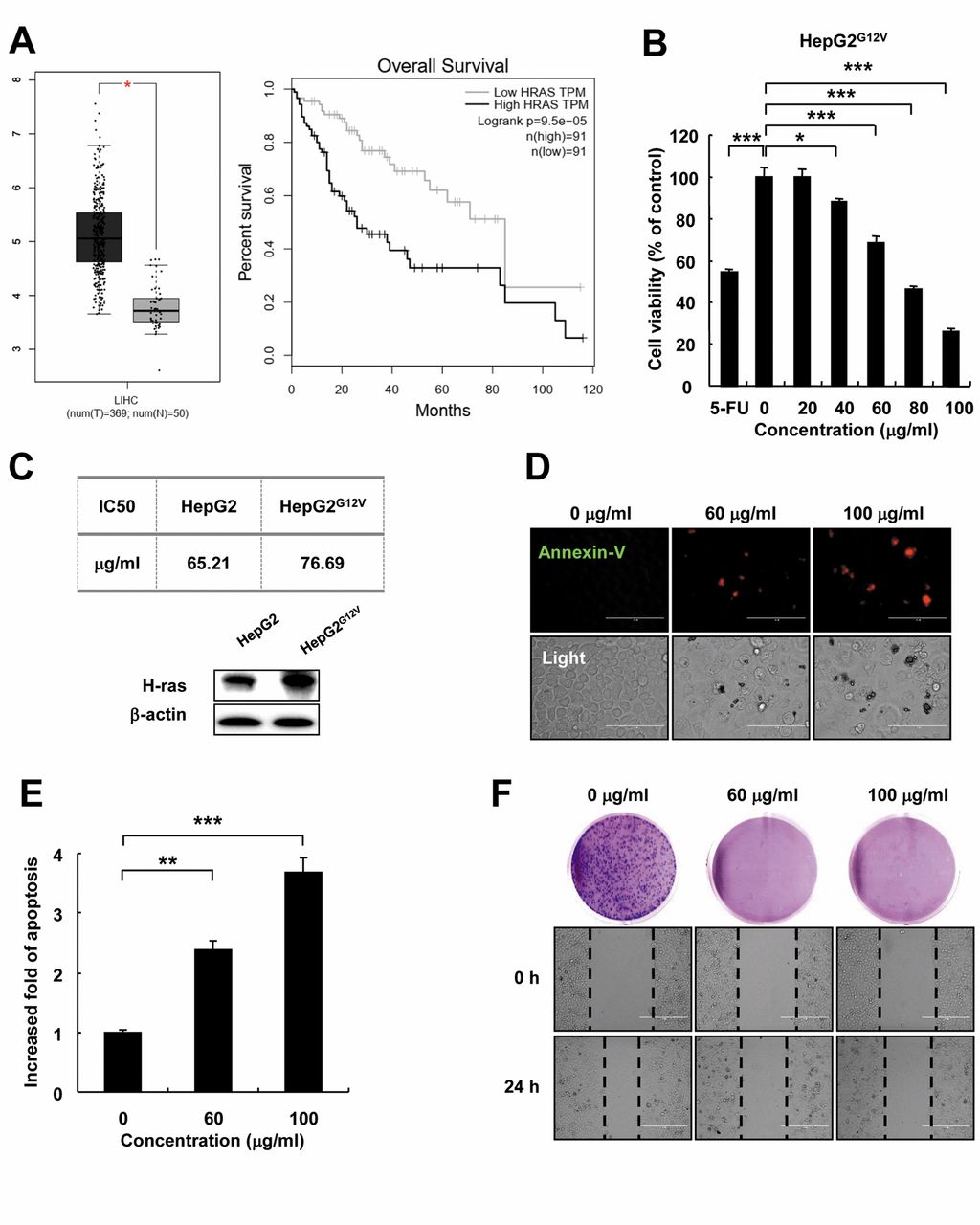
Effect of PQ on apoptosis, migration and colony formation ability in HepG2G12V cells. (A) The expression level of H-Ras in tumor and normal patients' liver tissue (dark gray represents high expression level of H-Ras in the liver tissue of a cancer patient, light gray corresponds to the H-Ras expression level in the liver tissue of a healthy person), *p<0.01; The survival rate analysis comparing patients with high and low expression of H-Ras in liver tissues (Black line represents survival in hepatoma patients with high expression level of H-Ras, light gray line represents survival in hepatoma patients with low expression level of H-Ras). (B) Effect of PQ treatment on cells viability detected by MTT Assay; (C) The IC50 values of PQ treated HepG2 cells and HepG2G12V cells were detected by MTT Assay, and the H-Ras expression in these two cell lines was detected by Western blot analysis; (D) Effect of PQ on HepG2G12V cells apoptosis. Annexin V-APC (Red) and bright field images (Light) were acquired with fluorescence microscope (scale bar=400 μm); (E) Quantitative analysis of figure D; (F) Effect of PQ on colony formation and migration ability in HepG2G12V cells (scale bar=100 μm). Data are presented as the mean±standard error of the mean of three different samples. (*p<0.05, **p<0.01, ***p<0.001).
PQ induces apoptosis through mitochondria dependent and MAPK signaling pathways in HepG2G12V cells. Mitogen-activated protein kinases (MAPKs) signals have many functions which regulate cell functions ranging from cell survival to cell death. It is closely related to cell apoptosis. Hence, the HepG2G12V cells were treated at different times (0, 12, 30, 60, 90, 120 min) with PQ (100 μg/ml) and p-Erk, Erk, p-P38, P38, p-Jnk and Jnk proteins of the MAPK family were detected. The results showed that P38 phosphorylation levels were increased while Erk and Jnk phosphorylation levels were decreased in response to PQ treatment in a time-dependent manner. Furthermore, the MAPK signaling pathway was also activated (Figure 3A). Prx family has the function of scavenging ROS. Subsequently, the expression levels of Prx family proteins (Prx I-VI) were detected after PQ treatment in different time points. Obviously, it shows that there was no obvious change in Prx I-V; however, the expression level of Prx VI was positively correlated with PQ treatment in a concentration-dependent manner (Figure 3B).
Inhibition of intracellular ROS reduces mitochondrial damage of HepG2G12V cells. Next, we investigated ROS accumulation as the key to PQ-induced apoptosis in HepG2G12V cells. To detect the levels of ROS, mitochondria damage and expression level of mitochondrial dependent apoptosis proteins in HepG2G12V cells, we treated cells with NAC (ROS inhibitors, 5 mM, Solarbio life sciences, Beijing, PR China) for 30 min followed by PQ treatment (100 μg/ml) for 24 h. The results showed that pretreatment with NAC could decrease intracellular ROS significantly (Figure 4A). Next, it was found that after NAC pre-treatment, MMP was decreased (Figure 4B). Concurrently, the level of ROS and synthesis of ATP in mitochondria was increased again (Figure 4C, D). In addition, the expression levels of apoptosis-related proteins, such as cleaved-caspase-3, cleaved-caspase-9 and BAD were decreased, while Bcl-xL levels were increased after NAC pre-treatment. Furthermore, the expression of pro-caspase-3 was decreased. However, the expression level of cleaved-caspase-8 was not changed in response to PQ treatment. Meanwhile, the expression level of H-Ras was increased (Figure 4E, F).
Inhibition of intracellular ROS reduces cellular apoptosis, cell migration and colony formation in HepG2G12V cells. To determine the correlation between PQ-induced apoptosis and ROS in HepG2G12V cells, apoptosis levels were detected after NAC pretreatment. The results showed that cells apoptosis was decreased after pre-treatment with NAC (Figure 5A). Simultaneously, the cell migration ability and colony formation ability in the NAC pretreatment group was increased, in comparison to the PQ treatment group (Figure 5B, C, D).
Discussion
In recent years, mitochondrial function has become one of the main research directions in cancer treatment due to the participation of mitochondria in various pathways in redox, cell metabolism and cell death (25-27). Mitochondria are the main source of intracellular reactive oxygen species. ROS are the by-products of mitochondrial electron transport chain reactions in cell respiration, including oxygen, hydroxyl radical (• OH), superoxide anion (O2−), singlet oxygen (O2) and hydrogen peroxide (H2O2) species. ROS plays an important role in cell growth, differentiation, metabolism, apoptosis and other signaling pathways (28, 29).
In the treatment of cancer, more and more drugs are aimed at regulating changes in ROS levels so as to induce apoptosis of cancer cells (30). In this study, we found that PQ can effectively increase the level of ROS both intracellularly and in mitochondria, and induce cell apoptosis. When cellular ROS levels increase rapidly, the permeability of the mitochondrial membrane changes and this affects the transmission of the mitochondrial respiratory chain (Figure 2A, B, C). This results in the inhibition of ATP synthesis (Figure 2D) and can lead to misregulation of cell metabolism. In this study, we found that after PQ treatment of HepG2G12V cells, changes of mitochondrial membrane permeability (MMP) and ATP synthesis were detected. In other studies, such as treatment of HepG2 cells with Brusatol, an increase in intracellular ROS has been reported, causing mitochondrial dysfunction, a reduction in ATP synthesis, and release of pro-apoptotic molecules to cause apoptosis (31). This is consistent with our findings, but whether PQ can affect other functions of HepG2G12V cells was unknown.

PQ induced ROS accumulation, mitochondrial dysfunction and apoptosis-associated protein expression changes in HepG2G12V cells. (A) Effect of PQ on cellular HepG2G12V ROS levels (fluorescence microscope images show the DHE in red, scale bar=400 μm); (B) Mitochondrial Membrane Potential Assay Kit with JC-1 staining was used to detect the changes in mitochondrial membrane potential in HepG2G12V cells after PQ treatment. The JC-1 detection showed a reduction in red fluorescence signal in response to PQ treatment (scale bar=100 μm); (C) Effect of PQ treatment on ROS level in mitochondria in HepG2G12V cells. MitoSOX (Red), DAPI (Blue) and bright field images (Light) were acquired in a fluorescence microscope (scale bar=100 μm); (D) ATP assay kit was used to detect the level of ATP production after PQ treatment. Data are presented as the mean±standard error of the mean of three different samples. *p<0.05; (E) Effect on c-caspase-8, c-caspase-9, pro-caspase-3 and c-caspase-3 proteins expression in HepG2G12V cells after treatment periods of 0, 6, 12 and 24 h with the 100 μg/ml of PQ; (F) Effects on H-Ras, Bcl-xL, and Bad proteins expression in HepG2G12V cells after treatment periods of 0, 6, 12 and 24 h with 100 μg/ml of PQ. Data are presented as the mean±standard error of the mean of three different samples.

PQ induces apoptosis through mitochondria-dependent and MAPK signaling activated pathways in HepG2G12V cells. (A) Effects on p-Erk, Erk, p-P38, P38, p-Jnk, Jnk and H-Ras expression in HepG2G12V cells after treatment periods of 0, 15, 30, 60, 90 and 120 min with 100 μg/ml of PQ. (B) Effects on Prx I-VI proteins expression in HepG2G12V cells after treatment periods of 0, 6, 12 and 24 h with 100 μg/ml of PQ. (C) Quantitative analysis of Prx I-VI proteins expression level. Data are presented as the mean standard error of the mean of three different samples. (*p<0.05, **p<0.01, ***p<0.001).
Apoptosis is induced by extrinsic and intrinsic pathways (32). The extrinsic pathway is activated by the death receptor-mediated caspase-8, activated caspase-8 acts as a signal to activate the downstream caspase-3 and induce apoptosis (33). The other pathway is the intrinsic pathway in which the damaged mitochondria release cytochrome c and activated caspase-9. The cleaved caspase-9 cleaves the caspase-3 and induces cell apoptosis (34). Bcl-2 family plays an important role in the regulation of mitochondrial permeability to induce apoptosis. There are pro-apoptotic proteins and anti-apoptotic proteins in the Bcl-2 family. When mitochondria are damaged, the pro-apoptotic proteins will release pro-apoptotic regulators into the cytoplasm; and the anti-apoptotic proteins will prevent the mitochondrial isolation from promoting apoptosis, together playing a role in regulating the mitochondrial-mediated apoptosis process (35-37).
When MMP is attacked by ROS and altered as a result, mitochondria release cytochrome c into the cytoplasm, activating the downstream caspase-9, which in turn activates caspase-3 cleavage, leading to apoptosis. We used Western Blot analysis, after PQ treatment of HepG2G12V cells, and there was no significant change in caspase-8, but caspase-9 was activated to cause the increase in caspase-3 cleavage. At the same time, the expression of pro-apoptotic protein BAD increased and the anti-apoptotic protein Bcl-xL was decreased, which shows that PQ-induced apoptosis of HepG2G12V cells may be induced by the mitochondrial intrinsic pathways. In order to determine the key role of ROS in PQ-induced apoptosis, we first detected the MMP and ATP synthesis after adding NAC (inhibitor of ROS). We found that after inhibiting ROS produced in cells, MMP damage and ATP synthesis were significantly reduced. The detection at the protein level also proved that the addition of NAC for pre-treatment reduced the activation of caspase-9, caspase-3 and BAD, and increased the expression level of Bcl-xL, while there was no change in caspase-8 expression. Furthermore, apoptosis was significantly reduced while cell migration and colony formation were also affected. These results indicate that ROS may be a key signal in PQ-induced HepG2G12V cell apoptosis.

Inhibition of intracellular ROS reduces mitochondrial damage and changes the expression of apoptotic protein in HepG2G12V cells. The HepG2G12V cells were pretreated with NAC for 30 min, followed with PQ treatment for 24 h. (A) The cellular ROS levels were visualized under a fluorescence microscope. (B) Damage of mitochondrial membrane potential was detected under fluorescent microscopy. Cell aggregates (Red fluorescence) and cell morphology (light) were visualized in a fluorescence bright field microscope. (C) Levels of ROS in mitochondria were detected under fluorescence microscopy. MitoSOX (Red), DAPI (Blue) and Light images were acquired in a fluorescence microscope. (D) Detection of ATP production in cells. *p<0.05; (E) The expression of proteins associated with mitochondria-dependent apoptosis protein were detected. (F) The expression of Bcl-xL, Bad, H-Ras protein were detected. Data are presented as the mean±standard error of the mean of three different samples.

Inhibition of intracellular ROS reduces cellular apoptosis, cell migration and colony formation in HepG2G12V cells. (A) The HepG2G12V cells were pretreated with NAC for 30 min, followed with PQ treatment for 24 h. The apoptosis in HepG2G12V cells is shown with fluorescence for Annexin V (red), upper panel, and light microscopy, lower panel, and images. (B) The HepG2G12V cells were pretreated with NAC for 30 min, followed with PQ treatment for 24 h. Detection of cell colony formation and cell migration ability. (C) Quantitative analysis of colony formation assay. (D) Quantitative analysis of cell migration. Data are presented as the mean±standard error of the mean of three different samples. (*p<0.05, ***p<0.001).
In the body, the production of ROS within the cell and the antioxidant system that maintains the steady state maintain a certain balance (38). When cell mitochondria are damaged, it causes an increase in the oxidative stress response. The most obvious characteristic is the increasing levels of ROS. At that moment, various oxidative balance systems in the body are adjusted, including peroxiredoxins (Prxs), superoxide dismutase (SOD), catalase (CAT), thioredoxin (TRX) and glutathione system. Previously in our lab, we mainly studied the role of Prx family in cancer cells. In order to detect the changes of antioxidant enzymes in the cells when the cells were stimulated by high levels of ROS, we examined the changes in the Peroxiredoxin family (Prxs) after PQ treatment. The Prxs are the main class of peroxidase in mammalian cells (39) and have the role of scavenging intracellular ROS (40). Our previous results have shown that Prx I can effectively regulate H-Ras-induced liver cancer apoptosis (41) while Prx II and Prx V can play a protective role in glutamate-induced HT22 apoptosis (42, 43) and the increase of Prx V can weaken β-lapachone or shikonin-induced SW480 apoptosis (44, 45). These results suggest that Prxs may have some regulatory effect on ROS-induced apoptosis. Our results show that after PQ treatment, the protein expression of Prx I-V slightly changed, but the protein expression of Prx VI showed a significant time-dependent change. This suggests that Prx VI may be the only Prxs that plays a regulatory role in PQ-induced HepG2G12V cell apoptosis.
The Ras/MAPK signal pathway is a classical signaling pathway and has been studied in Ras-induced mutations (46). In our results, we examined the changes in the MAPK pathway within a short period after PQ-treatment and found that with drug treatment, this pathway was activated and the level of H-Ras protein expression decreased. This indicates that Ras/MAPK signaling is involved in the PQ-induced HepG2G12V cell apoptosis pathway.
In this study, PQ caused cell apoptosis by inducing increased intracellular and mitochondrial ROS levels, triggering cell mitochondrial damage, and reduced ATP synthesis and also inhibited cell migration and colony formation in HepG2G12V cells. The key to apoptosis caused by PQ may be caused by the mitochondrial-dependent intrinsic apoptosis pathway. When it inhibited the production of ROS, the results were contrary to the previous results, which further proved the importance of ROS in PQ induced apoptosis of HepG2G12V cells. PQ can reduce the expression level of H-Ras in liver cancer cells and thus reduce the abnormal proliferation of cells, and also increase the expression change of Prx-VI to regulate the redox signal in these cells. Our results provide the new ideas and references for PQ to treat HepG2G12V liver cancer.
Acknowledgements
This study was supported by grants from the Korean Research Institute of Bioscience and Biotechnology Research Initiative Program (KRIBB) (KGM5162021). This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2017R1D1A1B03028188, 2020R1I1A2052417).
Footnotes
↵# These Authors contributed equally to this work.
Authors' Contributions
DPX, YXG, YHJ, TK and HNS completed experiment and wrote the manuscript. CXR, YL, YHH, MHJ, DZ, QZP, LYY, DSL and JYH performed data analysis, JK, JL and YHP provided P. quassioides extract reagents, TK, YDC and HNS had substantial contributions to conception and design. All Authors read and approved the final manuscript.
Conflicts of Interest
The Authors have no conflicts of interest to declare.
Funding
This research was supported by the project for postgraduate to innovate scientific research in Heilongjiang Bayi Agricultural University (No: YJSCX2019-Y64) and the scientific research team support plan of Heilongjiang Bayi Agricultural University (TDJH201904), PR China.
- Received May 18, 2020.
- Revision received June 1, 2020.
- Accepted June 6, 2020.
- Copyright© 2020, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}