

Abstract
Small heat shock proteins (small Hsps) are stress-induced molecular chaperones that act as holdases towards polypeptides that have lost their folding in stress conditions or consequently of mutations in their coding sequence. A cellular protection against the deleterious effects mediated by damaged proteins is thus provided to cells. These chaperones are also highly expressed in response to protein conformational and inflammatory diseases and cancer pathologies. Through specific and reversible modifications in their phospho-oligomeric organization, small Hsps can chaperone appropriate client proteins in order to provide cells with resistance to different types of injuries or pathological conditions. By helping cells to better cope with their pathological status, their expression can be either beneficial, such as in diseases characterized by pathological cell degeneration, or deleterious when they are required for tumor cell survival. Moreover, small Hsps are actively released by cells and can act as immunogenic molecules that have dual effects depending on the pathology. The cellular consequences linked to their expression levels and relationships with other Hsps as well as therapeutic strategies are discussed in view of their dynamic structural organization required to interact with specific client polypeptides.
1. Introduction
In the early sixties, Ritossa published papers reporting that the pattern of puffing in Drosophila chromosomes was drastically altered when third instar larvae were exposed to sublethal temperatures (35°C) or to the metabolic uncoupler dinitrophenol [1, 2]. This discovery, in addition of being the first illustration that environmental changes could modify the structure of chromosomes, suggested that new RNA messengers encoding polypeptides were synthesized in response to insults. Ten years later, these proteins were identified by Tissiéres et al. [3] and called heat shock proteins (Hsps). Thereafter, this cellular response was shown to be conserved from bacteria to human, including plants, and to be triggered by many environmental stress conditions such as starvation, exercise, recovery from hypoxia, infection, UV light, inflammation and nitrogen deficiency as well as toxins (arsenic, alcohols, metals, metabolic uncouplers, anticancer drugs, and many others). This led to the conclusion that a strong positive correlation exists between the presence of heat shock proteins and the ability of organisms to withstand stress and to transiently develop resistance [4–7]. In view of these observations, Hsps were also referred to as stress proteins, and their expression is now part of the so-called cellular stress response [7]. Five families of Hsps are induced by stress: the 70 kDa (HspA-Hsp70) family, the 20–30 kDa (HspB-small Hsps, sHsps) family, the 90 kDa (HspC-Hsp90) family, the 60 kDa (HspD-Hsp60) family, and the HspH (large Hsps) family [8]. Studies were then oriented to respond to two major questions: what is the mechanism of induction of Hsps and what is their role in the stressed cell? Stress-induced transcription of Hsps genes was rapidly found to depend on the activation of a particular transcription factor called heat shock factor 1 (HSF1). Indeed, following posttranslational modifications and homotrimer formation [9, 10], cytoplasmic HSF1 is activated [11] and migrates into the nucleus to induce a massive transcription of Hsp genes [12, 13]. Towards the second question, investigators discovered that the common denominator to the different conditions and agents that induce the expression of Hsps was their ability to alter the folding of proteins, particularly newly synthesized polypeptides that are in the process of being folded [6, 14, 15]. On a more general point of view, Hsps are expressed when the cellular environment becomes deleterious and disturbs the tertiary structure of polypeptides. So, numerous conditions and agents can induce Hsps synthesis. It was then shown that Hsps are molecular chaperones [16–18] that attenuate protein folding alterations during stress and allow amplified levels of repair and refolding of damaged polypeptides during stress recovery [6, 7]. Hence, Hsps protect proteins and help them to regain a functional tertiary structure without inducing any structural alterations. The next finding was the intriguing observation that Hsps are also constitutively expressed, that is, in the absence of apparent stress conditions (as, e.g., during cell growth, differentiation, and aging), and can act as specialized chaperones in different molecular mechanisms, such as those regulating intracellular transport, cytoskeleton architecture, intracellular redox status, stabilization of specific polypeptides, and protection against spontaneous or stimulated cell death [19]. Moreover, as described below, high levels of Hsps expression is common to many pathological conditions. Taken together, these facts open a road for new medical investigations leading to a recent explosive growth of the published studies dealing with heat shock proteins in human diseases.
Amongst Hsps, a subfamily of polypeptides in the 20–30 kDa range is characterized by the group of small stress proteins or small Hsps (HspB polypeptides) (Figure 1(a)). These proteins share a C-terminal domain in their sequence (about 40% of the proteins) which is also found in the major protein of mammalian crystallin: the alphaB-crystallin polypeptide [19–21], a less conserved N-terminal domain decorated with an hydrophobic WD/PF motif and phospho-serine sites [22], and a flexible C-terminal tail [23] containing a IXI/V motif [24]. Small Hsps also share the property to form large oligomeric structures (200–800 kDa) [19]. The human family of small Hsps contains ten members (HspB1 to HspB10) [25] plus the less conserved Hsp16.2 polypeptide [26] (see Figure 1(b)). Only four of them (HspB1, HspB5, HspB8, and HspB11) are induced by heat shock or other types of stress and five (HspB1, HspB4, HspB5, HspB8, and HspB11) bear a conserved ATP-independent chaperone activity [27, 28]. In this regard, up to now, the most studied chaperones have been HspB1 (also denoted Hsp27 or Hsp28) and the alphaA- and alphaB-crystallin polypeptides (HspB4 and HspB5). This paper discusses the multiple roles of these small Hsps in human diseases.
Figure 1.
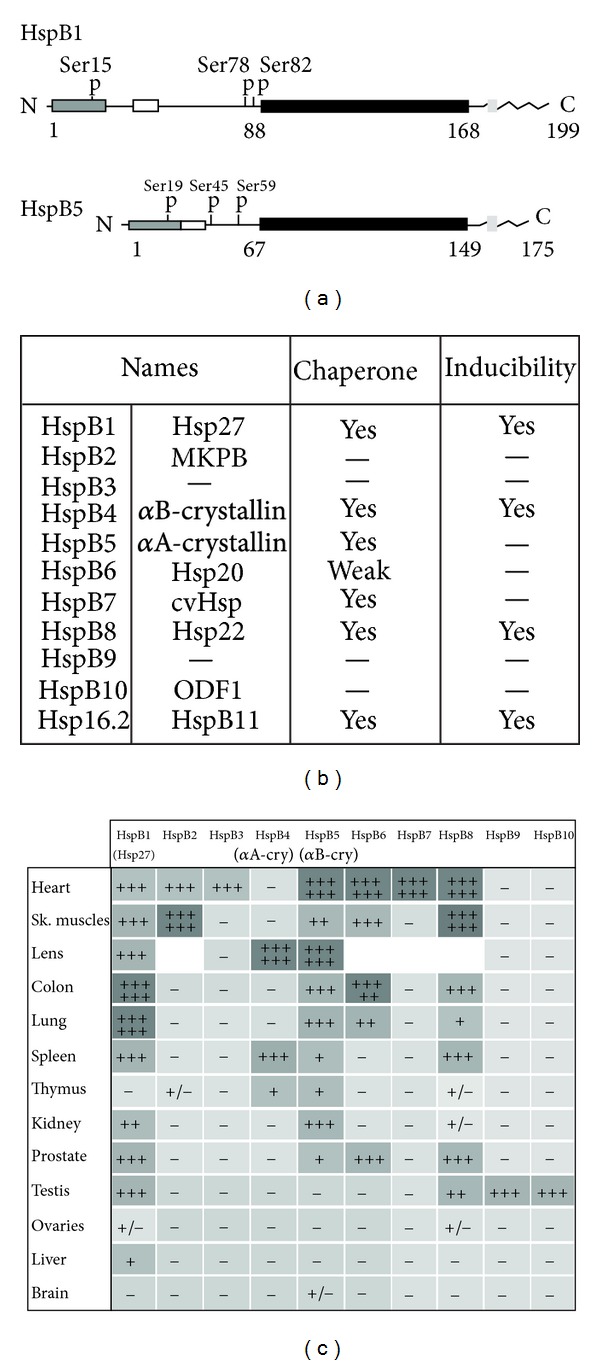
Small Hsps. (a) Organization of human HspB1 (Hsp27) and HspB5 (αB-crystallin) protein sequences. N-terminal gray box: WD/EPF domain; N-terminal white box: conserved sequence; black box: alpha crystallin domain; C-terminal gray box: IXI/V motif; the C-terminal flexible domain is also indicated. P: phosphorylated serine residues. Amino acids number is indicated. (b) Description of the members of the human small heat shock family of proteins. The distant Hsp16.2 polypeptide contains only a fraction of the alpha-crystallin domain. Chaperone activity and inducibility by heat shock are indicated. (c) Tissue-specific expression of constitutive small Hsps (compilation of murine as well as some human data).
2. Small Hsps Are Protective Molecular Chaperones towards Environmental Conditions or Agents That Alter Protein Conformation Homeostasis
As of today, the molecular function of several high molecular weight Hsps (Hsp70, Hsp90, Hsp60) is well documented (i.e., ATP-dependent chaperones), while that of the small Hsps was, until recently, more confuse in spite of the property of some of them to act as ATP-independent chaperones [29, 30]. In stress conditions, such as heat shock, small Hsps accumulate in order to trap and store stress-altered polypeptides in a refolding competent state that can interfere with their propensity to aggregate [26, 29, 31–34]. The name“holdase” has been proposed for this intriguing activity which depends on the dynamic oligomerization/phosphorylation status of small Hsps [30, 35–39]. Indeed, subsequently to stress-induced disruption of their oligomeric distribution, these Hsps interact with stress-altered polypeptides and store them via the reformation of large oligomeric structures [40–43]. By doing so, the large oligomeric complexes (up to 800 kDa, in case of HspB1) act as reservoirs that can further increase their sizes if more nonnative proteins accumulate. Stored polypeptides are in a folding competent state and can subsequently be refolded by the ATP-dependent “foldase” chaperone machines (Hsp70, Hsp90, and co-chaperones) [44–47] or degraded by the ubiquin-26S proteasome after being recognized by Hsp70 interacting E3 ubiquitin ligase CHIP [48]. Holdase and foldase machines are part of a coordinated network aimed at refolding or promoting the degradation of denatured polypeptides, a phenomenon which is essential for cell survival to acute stress. Small Hsps are cytoplasmic polypeptides, except in heat shock conditions, where some of them, such as HspB1, can be recovered in the nucleus at the level of granular structures [35] that have recently been shown to contain denatured proteins [49] that are stored for subsequent degradation during heat shock recovery [50]. In the nucleus of stressed myoblast cells, HspB1 as well as HspB5 also interact with intranuclear lamin to stabilize this stress-sensitive network [51]. In addition to the modulation of mRNA translation consequently of the trapping of eIF4G initiation translation factor in insoluble heat shock granules [52], a sumoylation-mediated feedback inhibition of HSF1 transactivation is another function of these proteins in response to heat shock [53].
HspB1 and HspB5 are very effective to protect cytoskeletal architecture homeostasis which is deeply altered in response to thermal or oxidative stress [54, 55]. In that respect, phosphorylated small HspB1 oligomers bear an F-actin capping activity that negatively modulates F-actin fibers growth and indirectly modulates extracellular matrix organization [56–58]. Consequently to its action towards F-actin, HspB1 indirectly regulates neutrophil chemotaxis and exocitosis, neurite outgrowth [59] and maintains sustained muscle contraction [60]. Moreover, in cancer cells, HspB1 is necessary for F-actin-mediated cytokinesis and therefore, interferes with the accumulation of giant polynucleated cells [61]. HspB1 and HspB5 also stabilize microtubules [62–64] while HspB5 has been described to be efficient towards intermediate filaments, particularly in muscle cells, where it associates with desmin [65, 66].
Another property of HspB1 and HspB5 is their ability to protect cells through an intriguing antioxidant property that decreases the levels of intracellular reactive oxygen species and nitric oxide and concomitantly upholds glutathione in its reducing form as well as mitochondrial membrane potential (ΔΦm) [30, 67–75]. Consequently, damages such as protein oxidation, lipid peroxidation, and cytoskeleton architecture disruption are attenuated [68–70]. Moreover, the positive effect towards ΔΦm provides the cell with an increased level of ATP production that stimulates the activity of ATP-dependent foldase chaperones.
To eliminate irreversibly damaged polypeptides, particularly the oxidized ones that cannot be refolded, HspB1 and HspB5 can trigger their degradation independently of the Hsp70-CHIP machine. Indeed, they can stimulate ubiquitination or, as in the case of HspB5, directly interact with the proteasome [76–79]. HspB8, which interacts with irreversibly altered proteins, can trigger the macroautophagy machinery, an ultimum mechanism to eliminate aggregated polypeptides generated by heat shock [80] or oxidative stress [81, 82], through a further association with Bag3 [83, 84].
3. Constitutively Expressed Small Hsps Maintain Protein Folding Homeostasis
Studies performed in different organisms have revealed an important property of small heat shock proteins, that is, their ability to be expressed in the absence of apparent stress in specific tissues of developing and adult organisms [85–90] (see Figure 1(c)). For example HspB1, which is highly abundant in muscles, is expressed in almost all tissues. In contrast, HspB4 (alphaA-crystallin) is almost exclusively present in lens cells while HspB5 (alphaB-crystallin), which associates with HspB4 to form the lens alpha-crystallin complex, is also constitutively expressed in tissues with high rates of oxidative metabolism, such as the heart, skeletal muscle fibers, brain, and kidney. The early phase of many differentiation processes is another example in which a high level of HspB1 is transiently expressed [91–98], and where this chaperone plays an essential role [99, 100]. One hypothesis could be that HspB1 secures differentiating cells from the toxicity of proteins that are of no more use or have generated inappropriate interactions. In that regard, HspB1 could participate in the mechanism that counteracts tendency of these proteins to form junk protein structures that could aggregate before they get degraded [97, 101]. On the other hand, it is not excluded that HspB1 could hold and protect essential polypeptides during the transient hostile intracellular environment of differentiating cells. As for example, cytoskeleton whose structure can be deeply modified during cell differentiation.
4. Small Hsps Are Beneficial in Protein Conformational and Inflammatory Diseases
Numerous studies have reported that elevated levels of constitutively expressed HspB1 and HspB5 are observed in pathological cells in which protein folding homeostasis is impaired by the accumulation of pathological proteins that are prone to aggregate, such as α-synuclein, β-amyloid peptide as well as polyQ mutants of huntingtin polypeptide that are responsive of Parkinson's, Alzheimer's and huntington, neurodegenerative diseases, respectively. HspB1 and/or HspB5 accumulate in cortical Lewy bodies, Alzheimer disease plaques, neurofibrillary tangles, Rosenthal fibers of Alexander's-disease, Creutzfeldt-Jakob altered neurones as well as in synuclein deposit associated to Parkinson's disease or myopathy-associated inclusion body [102–106]. HspB1 and HspB5 stimulate, through their holdase activity, the cellular resistance by attenuating aggregates formation, as for example, in myocardial infarction and cerebral ischemia [107, 108] (see Table 1). Consequently, they have been described as being able to promote cardioprotection [109] and to enhance nerve survival [110]. Similarly, overexpression of HspB6, HspB7, HspB8 as well as HspB1 can independently protect against tachycardia remodeling [111, 112]. By doing so, these proteins provide a beneficit that helps cells to counteract the development of pathological process that could lead to cardiomyopathic, neurodegenerative, myopathic, cataract, and retina diseases [73, 113–118]. The in vivo protective activity of these proteins as potent suppressors of cell degeneration was further confirmed in transgenic mice overexpressing HspB1 that are strongly protected against myocardial infarction and cerebral ischemia [107, 108]. These facts were also confirmed by the discovery of mutations in HspB1, HspB5, and HspB8 genes that inhibit their chaperone activity and provoke human diseases such as inherited peripheral and motor neuropathies, amyotrophic lateral sclerosis (ALS), axonal Charcot-Marie-Tooth disease, myofibrillar myopathies, cardiomyopathies, and cataracts [119–125]. The name αB-crystallinopathies has been given to the pathologies induced by mutations in HspB5. The protection may rely, at least in part, on the ability of these chaperones to specifically induce the sequestration of toxic protein oligomers [126]. Among the other members of the small Hsp family, HspB8 has the particular property to block polyglutamine (polyQ) huntingtin inclusion formation suggesting that it maintains aggregation prone polypeptides in a soluble state competent for rapid degradation. The C-terminal domain of HspB8 appears essential for this function [33]. HspB7 is even more potent since it not only suppresses polyQ aggregation, but also prevents polyQ-induced cellular toxicity; however, unlike HspB1, it does not improve the refolding of heat-denatured polypeptides [127].
Table 1.
Beneficial and deleterious roles of small Hsps. Schematic illustration of the dual role of intracellular and extracellular small Hsps as well as autoantibodies against these proteins.
| Beneficial role | Deleterious role | |
|---|---|---|
| Neurodegenerations | ||
| Myopathies | ||
| Cardioprotection | Intracellular small Hsps | |
| Cataracts | Cancers | |
| Strokes | ||
| Asthma | ||
| Inflammatory disease | ||
|
| ||
| Atherosclerosis Antiinflammatory effects Anticardiovascular effects |
Extracellular, circulating small Hsps | Tumor progression Metastasis Increased postinjuries infection Immunosuppression |
|
| ||
| Avoid propagation of insults | Facilitate retinal apoptosis | |
| Autoantibodies | Psoriasis | |
| Autoimmune diseases | ||
Through their ability to act as antioxidant molecules [67, 73, 74, 113, 128–130], HspB1 and HspB5 can be highly beneficial to cells expressing aggregated polypeptides. Indeed, oxidative stress is often a common feature of cells bearing aggregated polypeptides [131, 132] and in several of the above described diseases the production of abnormally high levels of deleterious intracellular reactive oxygen species has been detected [130, 132–138]. This is particularly the case in cells expressing pathological huntingtin, β-amyloid, or α-synuclein polypeptides which are iron/copper binding or metal homeostasis modulating polypeptides [139, 140] that can act as catalyzers and disregulate the hydroxyl radical generating Fenton reaction [141, 142]. Oxidative stress may then alter mitochondrial and proteasome function and aggravate protein aggregation [73, 113, 130, 143, 144].
Inflammatory pathologies, such as asthma, are other examples, where the antioxidant property of small Hsps has a beneficial protective role [145–149]. Indeed, through modulation of intracellular redox state and TAK-1 activity, these proteins interfere with tumor necrosis factor (TNFα) signaling pathways and therefore, negatively modulate inflammation processes [145, 147]. Moreover, HspB1 has been described to suppress skeletal muscle atrophy through its interaction with the activating kinases IKK-α and IKK-β of the transcription factor NF-κB [148]. One can also cite ischemic-related stroke injuries and alcoholic liver diseases characterized by the presence of Mallory bodies [108, 150]. These observations suggest crucial roles of HspB1 and HspB5 in inflammatory processes.
5. Small Hsps Are Antiapoptotic Proteins
Apoptosis, which differs from necrotic cell death, by being a genetically programmed process that requires energy, is negatively modulated by constitutively expressed Hsps. Indeed, in contrast to cells exposed to environmental insults, such as heat shock, where Hsps are synthesized to fight against the damaging effects of stress, no upregulation of Hsps expression occurs in cells committed to apoptosis. The reason is that a cell undergoing apoptosis does not fight against its own decision to commit suicide. The problem exists because Hsps, and particularly HspB1 and HspB5, are often constitutively expressed, particularly in human cancer cells, where they counteract an apoptotic process decided by the cell. In this type of cell death which does not induce the accumulation of misfolded polypeptides, HspB1 and HspB5 interact with specific protein _targets located along the signal transduction pathways activated by death receptors [151–155] as well as both upstream and downstream of mitochondria [156–162]. Complex and signal transduction-dependent structural reorganizations of HspB1 phosphorylation/oligomerization are observed in cells committed to apoptosis [39, 163] suggesting that this chaperone has multiple strategies to counteract apoptosis. Structural changes are probably needed to allow HspB1 to interact with specific _targets. Among them, one can cite: cytochrome c [157, 159], procaspase-3 [61, 152], Daxx [151], Stat3 [164], eIF4E [165], F-actin [159], HDAC6 [61], Stat2 [61], PTEN [166], and the cell survival kinase Akt [153, 155, 167, 168] that indirectly antagonizes Bax-mediated mitochondrial damages [162] and PEA-15-dependent Fas-induced apoptosis [169]. In addition to sharing some of HspB1 antiapoptotic mechanisms, such as caspase-3 maturation inhibition [154, 170, 171], HspB5 has specific ways to interfere with apoptosis. For example, it blocks the translocation to the mitochondria of the anti-apoptotic polypeptides Bax and Bcl-xs [160] and inhibits the activation of the proto-oncogene RAS [161]. Both HspB5 and HspB4 also modulate Akt, PKCα, and Raf/MEK/ERK pathways [172]. However, it cannot be concluded that all small Hsps are anti-apoptotic proteins per se since in some circumstances they can have the reverse effect: for example, HspB5 phosphorylated at the level of serine59 is proapoptotic since it prevents Bcl-2 translocation to mitochondria [173]. Moreover, depending on the cell type, HspB8 has pro- or anti-apoptotic activity.
6. Deleterious Effect Mediated by Small Hsps Expression in Human Cancer Pathologies
Many cancer cells express high loads of Hsps, such as HspB1 and HspB5; a phenomenon which increases their resistance to numerous deleterious agents and conditions [174–177] (see Table 1). One attractive, but still not proven, mechanism to explain a phenomenon linked to increased levels of HSF1 expression [178] is the “addiction to chaperones” hypothesis [177, 179]. Addiction could be caused by profound alterations in protein homeostasis resulting from mutant proteins that accumulate in cancer cells [178, 180]. So, contrasting to their beneficial role in degenerative and inflamatory diseases, their ability to protect cancer cells could be highly deleterious on a patient point of view. In that respect, HspB1 and HspB5 are essential for the growth of cancer cells and protect them against apoptotic or other types of death triggered by the immune system in the aim of their elimination [154, 169, 181–184]. They also provide cancer cells with the ability to counteract host anticancer response, such as senescence. This leads to aggressive cell growth [185, 186], metastasis formation, dissemination [187–190], and poor prognosis [174, 175]. Many studies have tried to decipher the mechanism that allow HspB1 to trigger tumor progression and metastasis. In that regard, several observations have already been made. For example, HspB1 can indirectly modulate extracellular matrix organization [56–58] through the stimulation of metalloproteinase type 2, an enzyme that efficiently digests the matrix surrounding tumor masses [191]. In addition, it can modulate cadherin-catenin cell adhesion polypeptides consequently to its interaction with cytoplasmic β-catenin [192]. More recently, HspB1 has been proposed to participate in the maintenance of breast cancer stem cells through regulation of the epithelial to mesenchymal transition process [193].
Another worth noting negative point concerns HspB1 ability to provide cancer cells with resistance to many anti-cancer drugs, which in turn, unfortunately, stimulates HspB1 expression [194–198]. Hence, high levels of HspB1 expression correlate with a poor clinical outcome of gastric, uterine, breast, prostate, ovarian, and head/neck cancers as well as of tumors from the urinary and nervous systems.
Other members of the family, such as HspB4 and HspB5, are also deeply involved in cancer biology. The first intriguing observation concerns HspB5 expression which transforms immortalized human mammary epithelial cells that can, subsequently to their injection in nude mice, develop in invasive mammary carcinomas that have the same aspect as basal-like breast tumors. At the molecular level, it has been found that HspB5-mediated growth of human breast basal-like tumor cells is epidermal growth factor (EGF)- and anchorage-independent. It increases cell migration and invasion through a constitutive activation of the MAPK kinase/ERK (MEK/ERK) pathway [199]. Hence, in addition to its anti-apoptotic property, HspB5 has the surprising ability to behave as an oncoprotein and consequently breast tumors expressing high levels of this protein are linked to short patient survival [200]. Contrasting with these observations, HspB4 expression in pancreatic cancer is a negative regulator of tumor development that has a good prognosis value [201].
7. Dual Role of Extracellular sHsps
The function of heat shock proteins goes beyond their intracellular localization and chaperone role since an increasing number of studies have recently described that, under normal physiological and stress conditions, a fraction of the cellular content of several Hsps, including the small Hsps, is recovered into the extracellular space, where they activate signaling pathways [202, 203]. The phenomenon, which is not related to cell injury or necrotic events, suggests a novel role of Hsps as universal proinflammatory intercellular “danger” signalling molecules. Hence, the classical role(s) of these highly conserved and ubiquitously expressed families of polypeptides is actually critically reevaluated. In that regard, Hsp60 was the first heat shock protein reported to be outside cultured cells [204]. Then, several studies demonstrated that Hsp70 and Hsp60 were localized on the cell surface [205–207], released in the extracellular milieu [208–212], and detected in the serum of normal and stressed individuals, together with circulating antibodies against these proteins [205, 213–215]. The level of Hsps in the serum of human individuals is highly variable and depends on multiple factors such as exercise [216], psychological stress [217], and diseases [211, 218]. This discovery has opened new roads of investigation aimed at understanding the role played by extracellular Hsps. It was first concluded that extracellular Hsps have a wide variety of functions towards neighboring cells including the possibility of being a danger signal to the immune system [219]. For example, it has been shown that in the brain, Hsp70 is released from glial cells and can subsequently interact with neurons and stimulate their ability to cope with stressful conditions [212]. Extracellular Hsp70 has also been reported to reduce neuronal polyglutamine toxicity and aggregation [220] and to change behavior in rats [221]. Circulating Hsp70 levels also predict, and may attenuate, the development of atherosclerosis in subjects with established hypertension [222]. On the opposite, in patients with colorectal cancer without distant metastasis, serum level of Hsp70 is associated with high mortality [223]. Another aspect of Hsp70 and Hsp60 deals with their immunogenicity and ability to activate dendritic cells as well as the production and secretion of cytokines [222, 224, 225]. Moreover, stimulation of both innate and adaptive forms of antitumor immune responses can be achieved through tumor-derived extracellular Hsp70-, Hsp90-, and gp96-peptide complexes that bind receptors on antigen presenting cells (APCs) and deliver tumor-specific antigens to major histocompatibility complex (MHC) class I molecules on the surface of such cells [207, 226–228]. Such antigen cross-presentation interactions form the basis for the “Hsp-based anticancer vaccines technology” [174, 229, 230] whose potency depends on the ability of Hsps to chaperone tumor antigenic peptides that stimulate antitumor immune responses through Hsp receptors [226, 231, 232].
Small Hsps have often been described as membrane associated proteins [233–235], and several recent reports point to their presence in the extracellular milieu. However, it is not yet known whether they could, similarly to the high molecular weight Hsps, elicit an immune response aimed at killing cancer cells through their association with immunogenic peptides. Despite this point, several positive and negative (for a patient point of view) functions of these extracellular proteins have already been reported (see Table 1). One interesting example concerns the atheroprotective effect of circulating HspB1 [236]. This protein, which has been known for quite a while to be an estrogen receptor beta (ERbeta)-associated protein, was noted for its role as a biomarker for atherosclerosis. The key experiment was the crossing of transgenic mice overexpressing HspB1 with apoE−/− mice that develop atherosclerosis when fed a high-fat diet. This experiment revealed a reduction in atherosclerotic lesion area in apoE−/−-HspB1 mice compared to apoE−/− mice. An interesting point of the phenomenon was its estrogen receptor-beta dependence. Indeed, it occurred only in females, where it correlated with a 10-fold higher level of circulating HspB1 compared to males. Moreover, there was a remarkable inverse correlation between circulating HspB1 levels and intensity of the lesions area. The atheroprotective activity of HspB1 was further confirmed by the inhibition of macrophage acLDL uptake and competition for the scavenger receptor by exogenous HspB1 added to culture media as well as by the decreased release of the proinflammatory cytokine interleukin-1β (IL-1β) and the increased release of the anti-inflammatory cytokine interleukin-10 (IL-10). Hence, the ovarian hormones mediated atheroprotective activity of HspB1 appears to be a consequence of its ability to compete for the uptake of atherogenic lipids and cholesterol and to attenuate vascular inflammation [237]. Based on the strong experimental evidence that ovarian hormones have a favorable effect on vessel wall homeostasis, HspB1 can therefore, be considered as an interesting _target that leads to the development of therapeutic drugs that can be used in replacement of the unfavorable risk-benefit profile of estrogen in vascular diseases preventing therapy of postmenopausal women [238]. It is also well-known that, in men and women, HspB1 shows an attenuated expression in human coronary arteries as the extent of atherosclerosis progresses. Up-regulation of HspB1 blocks this progression as demonstrated in transgenic mice overexpressing this protein. In a mechanistical point of view, it has recently been reported that recombinant HspB1 added to macrophages activates NF-κB and consequently changes the balance in the expression of key pro- and anti-inflammatory cytokines and antagonists of inflammation. These HspB1 triggered NF-κB-dependent signalings may explain the favorable net effect of HspB1 on the vessel wall [239]. Another example deals with the cardiovasculature which is probably the most exposed body system to stress. Hsps in the heart are known to be cardioprotective and their secreted counterparts play essential roles in the function of the cardiovascular tissues. In that respect, a positive action of circulating HspB1 has been demonstrated which deals with its anti-inflammatory capability that attenuates cardiovascular pathology [240]. On the negative side, high levels of HspB1 cell surface expression correlates with tumor growth and ability to metastasize [241]. Moreover, high levels of circulating HspB1 are also associated with tumor progression and increased postinjury infection [242–244]. By altering monocyte-derived dentritic cells to mediate immunosuppression, extracellular HspB1 has been proposed to have immunoregulatory activities that could contribute to immunopathology. Several other examples exist concerning disease-induced changes in the level of circulating HspB1; however, it is still unknown whether the phenomenon can be beneficial or not for the patient. For example, increased levels of circulating HspB1 are associated with micro- and macrovascular complications in type 1 diabetic patients and considered as a novel marker for diabetic neuropathy [245]. Hence, circulating extracellular small Hsps can have pathology-dependent dual roles similarly to their intracellular counterparts. The role of these extracellular proteins in normal physiological conditions is still not known, and speculations are open.
Because Hsps are intracellular proteins, a mechanism for their release into extracellular space must exist but remains obscur. First, it should be noted that Hsps are devoid of secretion signals, and their release is not blocked by inhibitors of ER-Golgi pathway, such as brefeldin A. Two mechanisms can be considered as follows: passive release consequently to necrotic cell death, trauma, or infection with lytic viruses and nonclassical active release. In that respect, active release can be triggered by agents, such as proinflammatory cytokines [208]. Recent observations suggest that, at least in the case of Hsp70, insertion of this Hsp into the plasma membrane requires inverse evagination, and its release from the cell is in a membrane-associated form (i.e., exosome) [212, 246, 247]. More precisely, the mechanism may involve surface membrane lipid rafts and the shedding of exosomes vesicles containing cytoplasmic constituents [203, 212, 248]. Strikingly, the tumor exosome-associated form of Hsp70 appears drastically more active than the free recombinant Hsp70 to stimulate macrophages [247] and natural killer cells [249]. Concerning the small Hsps, an interesting observation has been made in breast cancer patients with lymph node metastases. These patients show increased levels of circulating HspB1-positive microparticles [250] as well as microparticles containing annexin V, Her2/neu, and BCRP1 (Breast Cancer Resistance Protein 1). The origin of these microparticles is unknown, but they could be exosomes, hence, suggesting that HspB1 is released from cancer cells by a mechanism close to that of Hsp70.
Concerning the _target receptors that are recognized by Hsps, many cell surface proteins have been described as possible candidates; however, they are characterized by low-affinity interactions with Hsps. Nevertheless, two groups have been defined that are weakly or indirectly recognized by Hsp70, Hsp60, and a member of the Hsp90 family, gp96: the Toll-like receptors (TLRs) and scavenger receptors (SRs) [251]. The TLRs are major pattern recognition receptors (PRRs). TLR2 and TLR4 are Hsp60, Hsp70, and gp96 receptors that activate NF-κB [252, 253]. In addition, CD14, a human monocyte cell surface polypeptide which couples LPS exposure to TLR4 activation, is also required for Hsp70-mediated induction of TNFα, IL-1β and IL-6 [254]. CD14 is also recognized by Rhizobium leguminosarum chaperonin Hsp60.3 to trigger cytokine production [255]. These observations further demonstrate that Hsps can have a dual role as chaperone and cytokine. SRs are receptors for chemically modified forms of lipoproteins, and some of them can interact at high affinity with Hsp70, Hsp60, gp96, and Hsp90 [256–258]. The effects mediated by these interactions are complex and can have opposite effects. For example, LOX-1 mediates Hsp70 immunogenicity and antigen presentation [256], while gp96 binding to SR-A1 is immunosuppressive [259]. Of interest, a recent report has linked the inhibition of immune antitumoral activity to exosomes bearing Hsp70 when they interact with Toll like-receptor-2 of myeloid-derived suppressive cells (MDSCs); a phenomenon which inhibits the development of antitumoral response [260]. Taken together, these observations point to the complexity of the role played by extracellular Hsps towards their already described receptors. Unfortunately, no cell surface polypeptides have yet been characterized as putative small Hsps cell surface receptors.
8. Circulating sHsps Autoantibodies
As mentioned above, fascinating observations have been made concerning circulating autoantibodies against Hsps which are detected under normal conditions but seem to be more abundant in response to environmental or occupational stress and in a number of diseases [261]. As immunodominant molecules, Hsps can stimulate the immune system, leading to the production of autoantibodies recognizing epitopes shared by microbial and human Hsps. Surprisingly, such antibodies can regulate the inflammatory response positively or negatively. One example concerns breast cancer cells which express elevated levels of Hsps, a phenomenon that quite often correlates with reduced survival. So, does this provoke a generalized immune response towards Hsps? The answer is no, since serum HspB1 and Hsp90 autoantibodies show elevated levels but not Hsp70 autoantibody. Moreover, contrasting with the reduced survival associated to Hsp90 antibody, antibody to HspB1 has the surprizing property to correlate with an improved rather than a reduced survival. This leads to the conclusion that high levels of Hsps in breast cancer cells do not provoke a generalized immune response, and that Hsps serum autoantibodies have distinct associations with survival [262]. Hence, levels of circulating Hsps and anti-Hsps antibodies are now considered as useful parameters in tumor diagnosis [174]. Another example, dealing with small Hsps, concerns the presence of antibodies to HspB1, HspB5, Hsp70, and vimentin in aqueous humor of patients suffering from retinal pathologies, such as normal tension glaucoma [263, 264]. Of particular interest was the observation that exogenously applied HspB1 antibody enters human retina neuronal cells through an endocytic mechanism. This inactivates intracellular HspB1 and subsequently facilitates neuronal apoptosis [265]. Hence, it is believed that autoantibodies to small Hsps may impair cell survival in selective diseases, particularly those related to the human eye [264, 265]. In addition, it has been proposed that HspB1 is a _target of the exaggerated T cell response in psoriasis and an antigenic link between psoriasis and inflammatory bowel disease, uveitis, or arteriosclerosis, which are clinically associated pathologies [266]. However, care should be taken before concluding that one fundamental property of small Hsps is to act as autoantigens. In that respect, an interesting example concerns HspB5 in multiple sclerosis [267]. In this pathology, HspB5 has been considered for many years as an autoantigen based on its effects on humoral and cellular responses. However, this statement is probably not correct since recent experiments have shown that HspB5, through its chaperone activity, can bind immunoglobulins with high affinity. This obviously refutes most of the serological data used to assign HspB5 as an autoantigen in multiple sclerosis [268].
Hence, extracellular Hsps and autoantibodies to Hsps are likely to act as indicators of the physiological conditions of cells. These factors can prime other cells, particularly those of the immune system, to avoid the propagation of the insult. The cellular communication mechanism for sensing extracellular Hsps has been called “the stress observation system” [203]. Depending on the pathology, this mechanism could obviously be beneficial or not to the patient.
9. Small Hsps Multiple Functions Result of Their Interactions with Client Polypeptides
Small Hsps are surprizing proteins that have an incredible number of unrelated cellular functions as illustrated by the effects associated to their over- or underexpression. This may result from small Hsps interactions with a large number of client proteins that are essential to many cellular processes. In that respect, the most studied protein is HspB1; a protein known to interact with up to 34 polypeptides [176, 177]. The phenomenon is reminiscent of the already described “Hsp90/client protein concept” [269, 270]. Hsp90 is known to interact with over 200 client polypeptides (for an updated list see: http://www.picard.ch/downl-loads) in order to modulate their activity and/or half life. Hence, similar to Hsp90, HspB1, and probably other small Hsps are global regulators of cell systems [271, 272]. Some of the major clients which need to interact with HspB1 to avoid proteolytic degradation are Her2 oncogene, procaspase 3, HDM2, the histone deacetylase HDAC6, the transcription factor Stat2, and PTEN [61, 166, 183, 197]. Amongst the many clients whose activity is modified by HspB1, one can cite the translation initiation factor 4E (eIF4E) which modulates the translational initiation process, a crucial parameter for cancer cell growth and proliferation [165]. HspB1 client proteins, essential in tumorigenic and metastatic process, are nowadays actively searched for.
How HspB1 recognizes client protein _targets? Based on what is known for Hsp90, whose interactions with cochaperones and clients occur through a variety of conformational states [273, 274], HspB1 may take advantage of its complex and dynamic oligomerization-phosphorylation properties to generate structural organizations that can interact with specific protein substrates [39, 61, 176, 177, 275]. In other words, it is now believed that HspB1 is an environmental sensor, which through specific changes in its apparent native size/phosphorylation can reprogram its pattern of interacting client protein _targets. Consequently, HspB1 dynamic interactome may allow cells to quickly respond and mount the more appropriate response to a particular condition or insult [39, 176]. How changes in cell physiology could modulate the structural organization of HspB1 is still an unsolved question. The phenomenon may rely, at least in part, on the complex patterns of MAPKAPK2,3-dependent phosphorylation of three serines sites located in the N-terminal domain of HspB1 [39, 176, 276, 277]. Unfortunately, no precise information is yet available concerning the structural organizations of HspB1 that recognize crucial client polypeptides. An increased complexity may arise in cells expressing several small Hsps. Indeed, these proteins can interact with each other to form multiple combinatorial oligomeric structures [278–281] that could bear new protein _targets recognition abilities.
10. Therapeutic Approaches
It is now well established that small heat shock proteins increase cellular resistance to damages induced by stress or pathological conditions. Hence, it would be interesting to stimulate their expression to protect cells that are suffering and dying because of pathological conditions, such as those encountered in protein conformational and inflammatory diseases. The aim of this approach, by using drugs that up-regulate small Hsps holdase activity in a definite tissue, is to strengthen the cellular homeostasis protein folding and redox status machineries. These are potent systems that exist in every cells but which are limited and can be overwhelmed by pathological polypeptides [282] or drastic oxidative conditions [74, 128]. Moreover, compounds able to boost the expression of single or multiple members of the HspB family have a cardioprotective role involved in the maintenance or restoration of tissue integrity and contractile function, probably through the important role played by these Hsps towards cardiac muscle cells [111, 112]. On the flip side, such an approach could be highly detrimental in case of pathologies, where Hsps are involved in the resistance of invading pathological cells that can kill the patient, such as cancer cells. Moreover, we do not know what could be the effects of such strategies towards circulating small Hsps. In spite of these limitations, efforts are nevertheless made to discover drugs that can specifically stimulate small Hsps expression in a define tissue. One interesting example concerns the beneficial protective effect of orally administered geranylgeranylacetone in transgenic mice suffering from HspB5 mutation-dependent cardiomyopathy [283]. The effect in the heart correlated with reduced amyloid aggregates and increased HspB1 and HspB8 expression. However, what could be the effect of geranylgeranylacetone in other pathologies and particularly in primary tumors? This point should be investigated. Mimicking the holdase activity of small Hsps by drugs or peptides is an other way to tackle the problem. For example, carnosine and its acetyl derivative are effective as anticataract drugs due to their chemical chaperone ability that mimics HspB4-HspB5 holdase activity [284, 285]. Peptide aptamers that interact with small Hsps and positively modulate their activity are also interesting towards degenerating diseases since they may lead to the generation of stimulating peptidomimetic drugs [286].
In cancer pathologies, the problem associated to small Hsps expression is far more complex than in protein conformational and inflammatory diseases. At first glance, the therapeutic strategies described above which consist in stimulating small Hsps expression and/or activity are not appropriate since they would result in an increased resistance and aggressivity of cancer cells. Moreover, what is the role of circulating small Hsps and of anti-small Hsps antibodies in cancer patients? Is it beneficial or deleterious? Do small Hsps, like several other Hsps, interact with cancer-specific antigenic peptides that stimulate both innate and adaptive forms of antitumor immune responses? If this is indeed the case, care will then have to be taken to choose strategies that do not disturb this particular activity when the protective one associated to intracellular small Hsps is inactivated.
Antisense DNA vectors [287] and more recently RNA interference (RNAi) technologies have been used to decrease the intracellular level of small Hsps and destabilize their interactome. In that respect, the most studied protein has been HspB1 whose decreased level sensitized cancer cells to apoptotic inducers, anticancer drugs, and radiations and reduced their tumorigenic potential [61, 188, 189, 196, 288, 289]. In tumor, this may lead in the degradation of HspB1 tumorigenic and metastatic client proteins. However, in other tissues, RNAi may also induce the depletion of useful proteins chaperoned by HspB1 and/or abolish HspB1 antiaggregation and antioxidative effects; phenomena that could generate pathological side effects or stimulate diseases.
The search for less broad and more specific ways to abolish or stimulate small Hsps activity is a very difficult task since it will have to modulate, in a definite cell type, the complex formed by the _targeted small Hsp with specific pathological clients or aggregated proteins. Moreover, these future procedures should not interfere with the activity of the _targeted small Hsp when it is expressed in other tissues or when it interact with other clients. In the meantime, a better knowledge of the holdase activity and structure of the different small Hsps present in human cells will be required to open the road to the search of drugs that could inhibit their interactions with specific clients. This is illustrated by a recent analysis of the architecture and dynamics of complexes formed in vitro between an oligomeric small Hsp and client which revealed that over 300 different stoichiometries of interaction are possible [290, 291]. The specificity of the interaction of small Hsps with clients has been confirmed by two recent studies. The first one dealt with two peptide aptamers that specifically recognize different molecular surfaces of HspB1 and attenuate its antiapoptotic, antitumorigenic and cytoprotective activities [275]. The second study concerns RP101 (Bromovinyldeoxyuridine, BVDU, Brivudine), an antiviral drug that improves the efficiency of human pancreatic cancer chemotherapy through interaction with two phenylalanine residues (Phe29 and Phe33) in the N-terminal domain of HspB1. RP101 inhibits HspB1 interaction with specific procancerous binding partners and stimulates caspases activation [292].
11. Conclusion
In the recent years reports dealing with the expression and involvement of small Hsps in human pathologies as diverse as neurodegeneration, myopathies, cardiomyopathies, cataracts, inflammatory diseases, and cancers have grown exponentially. Until recently, it was believed that these Hsps were specialized molecular chaperones mainly synthesized in stress conditions and whose activity was to attenuate the damages to cellular proteins by inducing their storage until they could be refolded. The recent findings clearly show that, together with other Hsps, these proteins can be constitutively expressed and have an incredible number of crucial roles in normal and pathological cells. These activities are probably linked to their abilities to recognize, interact, and modulate the activity and/or half-life of many specific protein client _targets. This particular protective role of small Hsps towards protein folding can have dual consequences: (i) by helping cells to better cope with their pathological status; they can be beneficial in diseases characterized by pathological cell degeneration, (ii) by helping cells that evade death and proliferate, such as cancer cells, the activity of small Hsps can be highly deleterious. A third consequence could be towards small Hsps that are actively released by cells. It is now well established that small Hsps are therapeutic _targets whose activity needs either to be stimulated or abolished depending on the pathology. To be efficient and propose strategies aimed at designing active molecules that could modulate the activity of these Hsps, future works will have to unravel the precise role of their multiple combinatorial phospho-oligomeric structures to understand their complex interactions with many specific client proteins. These studies together with structural work [293, 294] and analysis of the organization of these proteins in living cells [39] will probably allow the discovery of new drugs testable for their effectiveness in different pathologies. As described here, the use of broad drug screening or genetic techniques to invalidate the activity or expression of these proteins could appear efficient, but on the long term they may prove to be disappointing due to unsuspected side-effects. Indeed, we should keep in mind the unfortunate modest effects and lack of FDA recognition reported to date for the broad inhibitors of Hsp90 chaperone activity in most cancer clinical trials [295].
Acknowledgments
This work was supported by grants from the Association Française pour les Myopathies (AFM), the Région Rhône-Alpes and Retina France. No conflict of interests is directly relevant to the content of this paper. The author addresses his sincere thanks to Dr. Patrick Mehlen for his help and for having him welcomed as an Emeritius Professor in his laboratory and to Valerie Arrigo for the comments on the paper.
References
- 1.Ritossa FM. Experimental activation of specific loci in polytene chromosomes of Drosophila. Experimental Cell Research. 1964;35(3):601–607. doi: 10.1016/0014-4827(64)90147-8. [DOI] [PubMed] [Google Scholar]
- 2.Ritossa F. Discovery of the heat shock response. Cell Stress & Chaperones. 1996;1(2):97–98. doi: 10.1379/1466-1268(1996)001<0097:dothsr>2.3.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Tissiéres A, Mitchell HK, Tracy UM. Protein synthesis in salivary glands of Drosophila melanogaster: relation to chromosome puffs. Journal of Molecular Biology. 1974;84(3):389–398. doi: 10.1016/0022-2836(74)90447-1. [DOI] [PubMed] [Google Scholar]
- 4.Craig EA. The stress response: changes in eukaryotic gene expression in response to environmental stress. Science. 1985;230(4727):800–801. doi: 10.1126/science.230.4727.800-a. [DOI] [PubMed] [Google Scholar]
- 5.Craig EA. The heat shock response. CRC Critical Reviews in Biochemistry. 1985;18(3):239–280. doi: 10.3109/10409238509085135. [DOI] [PubMed] [Google Scholar]
- 6.Lindquist S, Craig EA. The heat-shock proteins. Annual Review of Genetics. 1988;22:631–677. doi: 10.1146/annurev.ge.22.120188.003215. [DOI] [PubMed] [Google Scholar]
- 7.de Maio A. Heat shock proteins: facts, thoughts, and dreams. Shock. 1999;11(1):1–12. doi: 10.1097/00024382-199901000-00001. [DOI] [PubMed] [Google Scholar]
- 8.Kampinga HH, Hageman J, Vos MJ, et al. Guidelines for the nomenclature of the human heat shock proteins. Cell Stress and Chaperones. 2009;14(1):105–111. doi: 10.1007/s12192-008-0068-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Sorger PK, Pelham HRB. Yeast heat shock factor is an essential DNA-binding protein that exhibits temperature-dependent phosphorylation. Cell. 1988;54(6):855–864. doi: 10.1016/s0092-8674(88)91219-6. [DOI] [PubMed] [Google Scholar]
- 10.Westwood JT, Wu C. Activation of Drosophila heat shock factor: conformational change associated with a monomer-to-trimer transition. Molecular and Cellular Biology. 1993;13(6):3481–3486. doi: 10.1128/mcb.13.6.3481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zou J, Guo Y, Guettouche T, Smith DF, Voellmy R. Repression of heat shock transcription factor HSF1 activation by HSP90 (HSP90 complex) that forms a stress-sensitive complex with HSF1. Cell. 1998;94(4):471–480. doi: 10.1016/s0092-8674(00)81588-3. [DOI] [PubMed] [Google Scholar]
- 12.Wu C. Heat shock transcription factors: structure and regulation. Annual Review of Cell and Developmental Biology. 1995;11:441–469. doi: 10.1146/annurev.cb.11.110195.002301. [DOI] [PubMed] [Google Scholar]
- 13.Calderwood SK, Xie Y, Wang X, Khaleque MA, Chou SD, et al. Signal transduction pathways leading to heat shock transcription. Signal Transduction Insights. 2010;2:13–24. doi: 10.4137/STI.S3994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zou J, Salminen WF, Roberts SM, Voellmy R. Correlation between glutathione oxidation and trimerization of heat shock factor 1, an early step in stress induction of the Hsp response. Cell Stress and Chaperones. 1998;3(2):130–141. doi: 10.1379/1466-1268(1998)003<0130:cbgoat>2.3.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ananthan J, Goldberg AL, Voellmy R. Abnormal proteins serve as eukaryotic stress signals and trigger the activation of heat shock genes. Science. 1986;232(4749):522–524. doi: 10.1126/science.3083508. [DOI] [PubMed] [Google Scholar]
- 16.Ellis RJ, Hemmingsen SM. Molecular chaperones: proteins essential for the biogenesis of some macromolecular structures. Trends in Biochemical Sciences. 1989;14(8):339–342. doi: 10.1016/0968-0004(89)90168-0. [DOI] [PubMed] [Google Scholar]
- 17.Ellis RJ, van der Vies SM, Hemmingsen SM. The molecular chaperone concept. Biochemical Society Symposia. 1989;55:145–153. [PubMed] [Google Scholar]
- 18.Ellis RJ. Molecular chaperones: assisting assembly in addition to folding. Trends in Biochemical Sciences. 2006;31(7):395–401. doi: 10.1016/j.tibs.2006.05.001. [DOI] [PubMed] [Google Scholar]
- 19.Arrigo AP, Landry J. Expression and function of the low-molecular-weight Heat shock proteins. In: Morimoto RI, Tissieres A, Georgopoulos C, editors. The Biology of Heat Shock Proteins and Molecular Chaperones. Cold Spring Harbor, NY, USA: Cold Spring Harbor Laboratory Press; 1994. pp. 335–373. [Google Scholar]
- 20.Ingolia TD, Craig EA. Four small Drosophila heat shock proteins are related to each other and mammalian a-crystallin. Proceedings of the National Academy of Sciences of the United States of America. 1982;79(7):2360–2364. doi: 10.1073/pnas.79.7.2360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.de Jong WW, Leunissen JAM, Voorter CEM. Evolution of the α-crystallin/small heat-shock protein family. Molecular Biology and Evolution. 1993;10(1):103–126. doi: 10.1093/oxfordjournals.molbev.a039992. [DOI] [PubMed] [Google Scholar]
- 22.Thériault JR, Lambert H, Chávez-Zobel AT, Charest G, Lavigne P, Landry J. Essential role of the NH2-terminal WD/EPF motif in the phosphorylation-activated protective function of mammalian Hsp27. Journal of Biological Chemistry. 2004;279(22):23463–23471. doi: 10.1074/jbc.M402325200. [DOI] [PubMed] [Google Scholar]
- 23.Takemoto L, Emmons T, Horwitz J. The C-terminal region of α-crystallin: involvement in protection against heat-induced denaturation. Biochemical Journal. 1993;294(2):435–438. doi: 10.1042/bj2940435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Pasta SY, Raman B, Ramakrishna T, Rao CM. The IXI/V motif in the C-terminal extension of α-crystallins: alternative interactions and oligomeric assemblies. Molecular Vision. 2004;10:655–662. [PubMed] [Google Scholar]
- 25.Kappe G, Franck E, Verschuure P, Boelens WC, Leunissen JA, et al. The human genome encodes 10 alpha-crystallin-related small heat shock proteins: HspB1-10. Cell Cell Stress and Chaperones. 2003;8(1):53–61. doi: 10.1379/1466-1268(2003)8<53:thgecs>2.0.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bellyei S, Szigeti A, Pozsgai E, et al. Preventing apoptotic cell death by a novel small heat shock protein. European Journal of Cell Biology. 2007;86(3):161–171. doi: 10.1016/j.ejcb.2006.12.004. [DOI] [PubMed] [Google Scholar]
- 27.Arrigo AP, Simon S, Gibert B, et al. Hsp27 (HspB1) and αB-crystallin (HspB5) as therapeutic _targets. FEBS Letters. 2007;581(19):3665–3674. doi: 10.1016/j.febslet.2007.04.033. [DOI] [PubMed] [Google Scholar]
- 28.Arrigo AP, Simon S. Expression and functions of heat shock proteins in the normal and pathological mammalian eye. Current Molecular Medicine. 2010;10(9):776–793. doi: 10.2174/156652410793937804. [DOI] [PubMed] [Google Scholar]
- 29.Jakob U, Gaestel M, Engel K, Buchner J. Small heat shock proteins are molecular chaperones. Journal of Biological Chemistry. 1993;268(3):1517–1520. [PubMed] [Google Scholar]
- 30.Rogalla T, Ehrnsperger M, Preville X, et al. Regulation of Hsp27 oligomerization, chaperone function, and protective activity against oxidative stress/tumor necrosis factor by phosphorylation. Journal of Biological Chemistry. 1999;274(27):18947–18956. doi: 10.1074/jbc.274.27.18947. [DOI] [PubMed] [Google Scholar]
- 31.Horwitz J, Huang QL, Ding LL. Alpha-crystallin can function as a molecular chaperone. Proceedings of the National Academy of Sciences of the United States. 1992;89:10449–10453. doi: 10.1073/pnas.89.21.10449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ganea E. Chaperone-like activity of α-crystallin and other small heat shock proteins. Current Protein and Peptide Science. 2001;2(3):205–225. doi: 10.2174/1389203013381107. [DOI] [PubMed] [Google Scholar]
- 33.Carra S, Sivilotti M, Zobel ATC, Lambert H, Landry J. HspB8, a small heat shock protein mutated in human neuromuscular disorders, has in vivo chaperone activity in cultured cells. Human Molecular Genetics. 2005;14(12):1659–1669. doi: 10.1093/hmg/ddi174. [DOI] [PubMed] [Google Scholar]
- 34.Markossian KA, Yudin IK, Kurganov BI. Mechanism of suppression of protein aggregation by α-crystallin. International Journal of Molecular Sciences. 2009;10(3):1314–1345. doi: 10.3390/ijms10031314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Arrigo AP, Suhan JP, Welch WJ. Dynamic changes in the structure and intracellular locale of the mammalian low-molecular-weight heat shock protein. Molecular and Cellular Biology. 1988;8(12):5059–5071. doi: 10.1128/mcb.8.12.5059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Préville X, Schultz H, Knauf U, Gaestel M, Arrigo A-P. Analysis of the role of Hsp25 phosphorylation reveals the importance of the oligomerization state of this small heat shock protein in its protective function against TNFα- and hydrogen peroxide-induced cell death. Journal of Cellular Biochemistry. 1998;69(4):436–452. [PubMed] [Google Scholar]
- 37.Lelj-Garolla B, Mauk AG. Self-association of a small heat shock protein. Journal of Molecular Biology. 2005;345(3):631–642. doi: 10.1016/j.jmb.2004.10.056. [DOI] [PubMed] [Google Scholar]
- 38.Lelj-Garolla B, Mauk AG. Self-association and chaperone activity of Hsp27 are thermally activated. Journal of Biological Chemistry. 2006;281(12):8169–8174. doi: 10.1074/jbc.M512553200. [DOI] [PubMed] [Google Scholar]
- 39.Paul C, Simon S, Gibert B, Virot S, Manero F, Arrigo AP. Dynamic processes that reflect anti-apoptotic strategies set up by HspB1 (Hsp27) Experimental Cell Research. 2010;316(9):1535–1552. doi: 10.1016/j.yexcr.2010.03.006. [DOI] [PubMed] [Google Scholar]
- 40.Lee GJ, Roseman AM, Saibil HR, Vierling E. A small heat shock protein stably binds heat-denatured model substrates and can maintain a substrate in a folding-competent state. The EMBO Journal. 1997;16(3):659–671. doi: 10.1093/emboj/16.3.659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ehrnsperger M, Gräber S, Gaestel M, Buchner J. Binding of non-native protein to Hsp25 during heat shock creates a reservoir of folding intermediates for reactivation. The EMBO Journal. 1997;16(2):221–229. doi: 10.1093/emboj/16.2.221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Ehrnsperger M, Gaestel M, Buchner J. Analysis of chaperone properties of small Hsp’s. Methods in Molecular Biology. 2000;99:421–429. doi: 10.1385/1-59259-054-3:421. [DOI] [PubMed] [Google Scholar]
- 43.Haslbeck M, Franzmann T, Weinfurtner D, Buchner J. Some like it hot: the structure and function of small heat-shock proteins. Nature Structural and Molecular Biology. 2005;12(10):842–846. doi: 10.1038/nsmb993. [DOI] [PubMed] [Google Scholar]
- 44.Freeman BC, Morimoto RI. The human cytological molecular chaperones hsp90, hsp70 (hsc70) and hdj-1 have distinct roles in recognition of a non-native protein and protein refolding. The EMBO Journal. 1996;15(12):2969–2979. [PMC free article] [PubMed] [Google Scholar]
- 45.Bukau B, Horwich AL. The Hsp70 and Hsp60 chaperone machines. Cell. 1998;92(3):351–366. doi: 10.1016/s0092-8674(00)80928-9. [DOI] [PubMed] [Google Scholar]
- 46.Buchner J. Hsp90 and Co.—a holding for folding. Trends in Biochemical Sciences. 1999;24(4):136–141. doi: 10.1016/s0968-0004(99)01373-0. [DOI] [PubMed] [Google Scholar]
- 47.Lee GJ, Vierling E. A small heat shock protein cooperates with heat shock protein 70 systems to reactivate a heat-denatured protein. Plant Physiology. 2000;122(1):189–198. doi: 10.1104/pp.122.1.189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.McDonough H, Patterson C. CHIP: a link between the chaperone and proteasome systems. Cell Stress and Chaperones. 2003;8(4):303–308. doi: 10.1379/1466-1268(2003)008<0303:calbtc>2.0.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Bryantsev AL, Loktionova SA, Ilyinskaya OP, Tararak EM, Kampinga HH, Kabakov AE. Distribution, phosphorylation, and activities of Hsp25 in heat-stressed H9c2 myoblasts: a functional link to cytoprotection. Cell Stress and Chaperones. 2002;7(2):146–155. doi: 10.1379/1466-1268(2002)007<0146:dpaaoh>2.0.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Bryantsev AL, Kurchashova SY, Golyshev SA, et al. Regulation of stress-induced intracellular sorting and chaperone function of Hsp27 (HspB1) in mammalian cells. Biochemical Journal. 2007;407(3):407–417. doi: 10.1042/BJ20070195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Adhikari AS, Sridhar Rao K, Rangaraj N, Parnaik VK, Mohan Rao C. Heat stress-induced localization of small heat shock proteins in mouse myoblasts: intranuclear lamin A/C speckles as _target for αB-crystallin and Hsp25. Experimental Cell Research. 2004;299(2):393–403. doi: 10.1016/j.yexcr.2004.05.032. [DOI] [PubMed] [Google Scholar]
- 52.Cuesta R, Laroia G, Schneider RJ. Chaperone Hsp27 inhibits translation during heat shock by binding eIF4G and facilitating dissociation of cap-initiation complexes. Genes and Development. 2000;14(12):1460–1470. [PMC free article] [PubMed] [Google Scholar]
- 53.Brunet Simioni M, de Thonel A, Hammann A, et al. Heat shock protein 27 is involved in SUMO-2/3 modification of heat shock factor 1 and thereby modulates the transcription factor activity. Oncogene. 2009;28(37):3332–3344. doi: 10.1038/onc.2009.188. [DOI] [PubMed] [Google Scholar]
- 54.Welch WJ, Feramisco JR. Disruption of the three cytoskeletal networks in mammalian cells does not affect transcription, translation, or protein translocation changes induced by heat shock. Molecular and Cellular Biology. 1985;5(7):1571–1581. doi: 10.1128/mcb.5.7.1571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Bellomo G, Mirabelli F. Oxidative stress and cytoskeletal alterations. Annals of the New York Academy of Sciences. 1992;663:97–109. doi: 10.1111/j.1749-6632.1992.tb38653.x. [DOI] [PubMed] [Google Scholar]
- 56.Perng MD, Cairns L, van den IJssel P, Prescott A, Hutcheson AM, Quinlan RA. Intermediate filament interactions can be altered by HSP27 and αB-crystallin. Journal of Cell Science. 1999;112, part 13:2099–2112. doi: 10.1242/jcs.112.13.2099. [DOI] [PubMed] [Google Scholar]
- 57.Dalle-Donne I, Rossi R, Milzani A, di Simplicio P, Colombo R. The actin cytoskeleton response to oxidants: from small heat shock protein phosphorylation to changes in the redox state of actin itself. Free Radical Biology and Medicine. 2001;31(12):1624–1632. doi: 10.1016/s0891-5849(01)00749-3. [DOI] [PubMed] [Google Scholar]
- 58.Mounier N, Arrigo A-P. Actin cytoskeleton and small heat shock proteins: How do they interact? Cell Stress and Chaperones. 2002;7(2):167–176. doi: 10.1379/1466-1268(2002)007<0167:acashs>2.0.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Jog NR, Jala VR, Ward RA, Rane MJ, Haribabu B, McLeish KR. Heat shock protein 27 regulates neutrophil chemotaxis and exocytosis through two independent mechanisms. Journal of Immunology. 2007;178(4):2421–2428. doi: 10.4049/jimmunol.178.4.2421. [DOI] [PubMed] [Google Scholar]
- 60.Yamboliev IA, Hedges JC, Mutnick JLM, Adam LP, Gerthoffer WT. Evidence for modulation of smooth muscle force by the p38 MAP kinase/HSP27 pathway. American Journal of Physiology—Heart and Circulatory Physiology. 2000;278(6):H1899–H1907. doi: 10.1152/ajpheart.2000.278.6.H1899. [DOI] [PubMed] [Google Scholar]
- 61.Gibert B, Eckel B, Fasquelle L, et al. Knock down of heat shock protein 27 (hspb1) induces degradation of several putative client proteins. PLoS One. 2012;7(1) doi: 10.1371/journal.pone.0029719.Article numbere29719 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Preville X, Mehlen P, Fabre-Jonca N, et al. Biochemical and immunofluorescence analysis of the constitutively expressed HSP27 stress protein in monkey CV-1 cells. Journal of Biosciences. 1996;21(2):221–234. [Google Scholar]
- 63.Hino M, Kurogi K, Okubo MA, Murata-Hori M, Hosoya H. Small heat shock protein 27 (HSP27) associates with tubulin/microtubules in HeLa cells. Biochemical and Biophysical Research Communications. 2000;271(1):164–169. doi: 10.1006/bbrc.2000.2553. [DOI] [PubMed] [Google Scholar]
- 64.Xi JH, Bai F, McGaha R, Andley UP. Alpha-crystallin expression affects microtubule assembly and prevents their aggregation. The FASEB Journal. 2006;20(7):846–857. doi: 10.1096/fj.05-5532com. [DOI] [PubMed] [Google Scholar]
- 65.Bennardini F, Wrzosek A, Chiesi M. αB-Crystallin in cardiac tissue: association with actin and desmin filaments. Circulation Research. 1992;71(2):288–294. doi: 10.1161/01.res.71.2.288. [DOI] [PubMed] [Google Scholar]
- 66.Djabali K, Piron G, de Néchaud B, Portier MM. αB-crystallin interacts with cytoplasmic intermediate filament bundles during mitosis. Experimental Cell Research. 1999;253(2):649–662. doi: 10.1006/excr.1999.4679. [DOI] [PubMed] [Google Scholar]
- 67.Mehlen P, Kretz-Remy C, Préville X, Arrigo AP. Human hsp27, Drosophila hsp27 and human αB-crystallin expression-mediated increase in glutathione is essential for the protective activity of these proteins against TNFα-induced cell death. The EMBO Journal. 1996;15(11):2695–2706. [PMC free article] [PubMed] [Google Scholar]
- 68.Préville X, Gaestel M, Arrigo A-P. Phosphorylation is not essential for protection of L929 cells by Hsp25 against H2O2-mediated disruption actin cytoskeleton, a protection which appears related to the redox change mediated by Hsp25. Cell Stress and Chaperones. 1998;3(3):177–187. doi: 10.1379/1466-1268(1998)003<0177:pinefp>2.3.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Préville X, Salvemini F, Giraud S, et al. Mammalian small stress proteins protect against oxidative stress through their ability to increase glucose-6-phosphate dehydrogenase activity and by maintaining optimal cellular detoxifying machinery. Experimental Cell Research. 1999;247(1):61–78. doi: 10.1006/excr.1998.4347. [DOI] [PubMed] [Google Scholar]
- 70.Paul C, Arrigo AP. Comparison of the protective activities generated by two survival proteins: Bcl-2 and Hsp27 in L929 murine fibroblasts exposed to menadione or staurosporine. Experimental Gerontology. 2000;35(6-7):757–766. doi: 10.1016/s0531-5565(00)00150-9. [DOI] [PubMed] [Google Scholar]
- 71.Arrigo AP. Hsp27: novel regulator of intracellular redox state. IUBMB Life. 2002;52(6):303–307. doi: 10.1080/152165401317291156. [DOI] [PubMed] [Google Scholar]
- 72.Yan LJ, Christians ES, Liu L, Xiao X, Sohal RS, Benjamin IJ. Mouse heat shock transcription factor 1 deficiency alters cardiac redox homeostasis and increases mitochondrial oxidative damage. The EMBO Journal. 2002;21(19):5164–5172. doi: 10.1093/emboj/cdf528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Firdaus WJJ, Wyttenbach A, Diaz-Latoud C, Currie RW, Arrigo AP. Analysis of oxidative events induced by expanded polyglutamine huntingtin exon 1 that are differentially restored by expression of heat shock proteins or treatment with an antioxidant. FEBS Journal. 2006;273(13):3076–3093. doi: 10.1111/j.1742-4658.2006.05318.x. [DOI] [PubMed] [Google Scholar]
- 74.Arrigo AP, Virot S, Chaufour S, Firdaus W, Kretz-Remy C, Diaz-Latoud C. Hsp27 consolidates intracellular redox homeostasis by upholding glutathione in its reduced form and by decreasing iron intracellular levels. Antioxidants and Redox Signaling. 2005;7(3-4):414–422. doi: 10.1089/ars.2005.7.414. [DOI] [PubMed] [Google Scholar]
- 75.Arrigo AP. The cellular “networking” of mammalian Hsp27 and its functions in the control of protein folding, redox state and apoptosis. Advances in Experimental Medicine and Biology. 2007;594:14–26. doi: 10.1007/978-0-387-39975-1_2. [DOI] [PubMed] [Google Scholar]
- 76.Boelens WC, Croes Y, de Jong WW. Interaction between αB-crystallin and the human 20S proteasomal subunit C8/α7. Biochimica et Biophysica Acta—Protein Structure and Molecular Enzymology. 2001;1544(1-2):311–319. doi: 10.1016/s0167-4838(00)00243-0. [DOI] [PubMed] [Google Scholar]
- 77.Parcellier A, Schmitt E, Gurbuxani S, et al. HSP27 is a ubiquitin-binding protein involved in I-κBα proteasomal degradation. Molecular and Cellular Biology. 2003;23(16):5790–5802. doi: 10.1128/MCB.23.16.5790-5802.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Den Engelsman J, Keijsers V, de Jong WW, Boelens WC. The small heat-shock protein αB-crystallin promotes FBX4-dependent ubiquitination. Journal of Biological Chemistry. 2003;278(7):4699–4704. doi: 10.1074/jbc.M211403200. [DOI] [PubMed] [Google Scholar]
- 79.Barbash O, Lin DI, Diehl JA. SCF Fbx4/alphaB-crystallin cyclin D1 ubiquitin ligase: a license to destroy. Cell Division. 2007;2, article 2 doi: 10.1186/1747-1028-2-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Nivon M, Richet E, Codogno P, Arrigo AP, Kretz-Remy C. Autophagy activation by NFκB is essential for cell survival after heat shock. Autophagy. 2009;5(6):766–783. doi: 10.4161/auto.8788. [DOI] [PubMed] [Google Scholar]
- 81.Keller JN, Dimayuga E, Chen Q, Thorpe J, Gee J, Ding Q. Autophagy, proteasomes, lipofuscin, and oxidative stress in the aging brain. International Journal of Biochemistry and Cell Biology. 2004;36(12):2376–2391. doi: 10.1016/j.biocel.2004.05.003. [DOI] [PubMed] [Google Scholar]
- 82.Kiffin R, Bandyopadhyay U, Cuervo AM. Oxidative stress and autophagy. Antioxidants and Redox Signaling. 2006;8(1-2):152–162. doi: 10.1089/ars.2006.8.152. [DOI] [PubMed] [Google Scholar]
- 83.Carra S, Seguin SJ, Landry J. HspB8 and Bag3: a new chaperone complex _targeting misfolded proteins to macroautophagy. Autophagy. 2008;4(2):237–239. doi: 10.4161/auto.5407. [DOI] [PubMed] [Google Scholar]
- 84.Carra S. The stress-inducible HspB8-Bag3 complex induces the eIF2α kinase pathway: implications for protein quality control and viral factory degradation? Autophagy. 2009;5(3):428–429. doi: 10.4161/auto.5.3.7894. [DOI] [PubMed] [Google Scholar]
- 85.Pauli D, Tonka CH, Tissieres A, Arrigo AP. Tissue-specific expression of the heat shock protein HSP27 during Drosophila melanogaster development. Journal of Cell Biology. 1990;111(3):817–828. doi: 10.1083/jcb.111.3.817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Bhat SP, Nagineni CN. αB subunit of lens-speciific protein α-crystallin is present in other ocular and non-ocular tissues. Biochemical and Biophysical Research Communications. 1989;158(1):319–325. doi: 10.1016/s0006-291x(89)80215-3. [DOI] [PubMed] [Google Scholar]
- 87.Srinivasan AN, Nagineni CN, Bhat SP. αA-crystallin is expressed in non-ocular tissues. Journal of Biological Chemistry. 1992;267(32):23337–23341. [PubMed] [Google Scholar]
- 88.Gernold M, Knauf U, Gaestel M, Stahl J, Kloetzel PM. Development and tissue-specific distribution of mouse small heat shock protein hsp25. Developmental Genetics. 1993;14(2):103–111. doi: 10.1002/dvg.1020140204. [DOI] [PubMed] [Google Scholar]
- 89.Huang L, Min JN, Masters S, Mivechi NF, Moskophidis D. Insights into function and regulation of small heat shock protein 25 (HSPB1) in a mouse model with _targeted gene disruption. Genesis. 2007;45(8):487–501. doi: 10.1002/dvg.20319. [DOI] [PubMed] [Google Scholar]
- 90.Tanguay RM, Wu Y, Khandjian EW. Tissue-specific expression of heat shock proteins of the mouse in the absence of stress. Developmental Genetics. 1993;14(2):112–118. doi: 10.1002/dvg.1020140205. [DOI] [PubMed] [Google Scholar]
- 91.Spector NL, Mehlen P, Ryan C, et al. Regulation of the 28 kDa heat shock protein by retinoic acid during differentiation of human leukemic HL-60 cells. FEBS Letters. 1994;337(2):184–188. doi: 10.1016/0014-5793(94)80270-x. [DOI] [PubMed] [Google Scholar]
- 92.Spector NL, Ryan C, Samson W, Levine H, Nadler LM, Arrigo AP. Heat shock protein is a unique marker of growth arrest during macrophage differentiation of HL-60 cells. Journal of Cellular Physiology. 1993;156(3):619–625. doi: 10.1002/jcp.1041560322. [DOI] [PubMed] [Google Scholar]
- 93.Chaufour S, Mehlen P, Arrigo AP. Transient accumulation, phosphorylation and changes in the oligomerization of Hsp27 during retinoic acid-induced differentiation of HL-60 cells: possible role in the control of cellular growth and differentiation. Cell Stress & Chaperones. 1996;1(4):225–235. doi: 10.1379/1466-1268(1996)001<0225:tapaci>2.3.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Davidson SM, Morange M. Hsp25 and the p38 MAPK pathway are involved in differentiation of cardiomyocytes. Developmental Biology. 2000;218(2):146–160. doi: 10.1006/dbio.1999.9596. [DOI] [PubMed] [Google Scholar]
- 95.Arrigo AP, Ducasse C. Expression of the anti-apoptotic protein Hsp27 during both the keratinocyte differentiation and dedifferentiation of HaCat cells: expression linked to changes in intracellular protein organization? Experimental Gerontology. 2002;37(10-11):1247–1255. doi: 10.1016/s0531-5565(02)00131-6. [DOI] [PubMed] [Google Scholar]
- 96.Duverger O, Paslaru L, Morange M. HSP25 is involved in two steps of the differentiation of PAM212 keratinocytes. Journal of Biological Chemistry. 2004;279(11):10252–10260. doi: 10.1074/jbc.M309906200. [DOI] [PubMed] [Google Scholar]
- 97.Arrigo AP. In search of the molecular mechanism by which small stress proteins counteract apoptosis during cellular differentiation. Journal of Cellular Biochemistry. 2005;94(2):241–246. doi: 10.1002/jcb.20349. [DOI] [PubMed] [Google Scholar]
- 98.Brown DD, Christine KS, Showell C, Conlon FL. Small heat shock protein Hsp27 is required for proper heart tube formation. Genesis. 2007;45(11):667–678. doi: 10.1002/dvg.20340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Mehlen P, Mehlen A, Godet J, Arrigo AP. Hsp27 as a switch between differentiation and apoptosis in murine embryonic stem cells. Journal of Biological Chemistry. 1997;272(50):31657–31665. doi: 10.1074/jbc.272.50.31657. [DOI] [PubMed] [Google Scholar]
- 100.Mehlen P, Coronas V, Ljubic-Thibal V, et al. Small stress protein Hsp27 accumulation during dopamine-mediated differentiation of rat olfactory neurons counteracts apoptosis. Cell Death and Differentiation. 1999;6(3):227–233. doi: 10.1038/sj.cdd.4400483. [DOI] [PubMed] [Google Scholar]
- 101.de Thonel A, Vandekerckhove J, Lanneau D, et al. HSP27 controls GATA-1 protein level during erythroid cell differentiation. Blood. 2010;116(1):85–96. doi: 10.1182/blood-2009-09-241778. [DOI] [PubMed] [Google Scholar]
- 102.Rankawek K, Bosman GJC, de Jong WW. Expression of small heat-shock protein hsp 27 in reactive gliosis in Alzheimer disease and other types of dementia. Acta Neuropathologica. 1994;87(5):511–519. doi: 10.1007/BF00294178. [DOI] [PubMed] [Google Scholar]
- 103.Muchowski PJ. Protein misfolding, amyloid formation, and neurodegeneration: a critical role for molecular chaperones? Neuron. 2002;35(1):9–12. doi: 10.1016/s0896-6273(02)00761-4. [DOI] [PubMed] [Google Scholar]
- 104.Wyttenbach A. Role of heat shock proteins during polyglutamine neurodegeneration: mechanisms and hypothesis. Journal of Molecular Neuroscience. 2004;23(1-2):69–95. doi: 10.1385/JMN:23:1-2:069. [DOI] [PubMed] [Google Scholar]
- 105.Muchowski PJ, Wacker JL. Modulation of neurodegeneration by molecular chaperones. Nature Reviews Neuroscience. 2005;6(1):11–22. doi: 10.1038/nrn1587. [DOI] [PubMed] [Google Scholar]
- 106.Bruinsma IB, Bruggink KA, Kinast K, et al. Inhibition of alpha-synuclein aggregation by small heat shock proteins. Proteins. 2011;79(10):2956–2967. doi: 10.1002/prot.23152. [DOI] [PubMed] [Google Scholar]
- 107.Latchman DS. Hsp27 and cell survival in neurones. International Journal of Hyperthermia. 2005;21(5):393–402. doi: 10.1080/02656730400023664. [DOI] [PubMed] [Google Scholar]
- 108.Efthymiou CA, Mocanu MM, de Belleroche J, Wells DJ, Latchmann DS, Yellon DM. Heat shock protein 27 protects the heart against myocardial infarction. Basic Research in Cardiology. 2004;99(6):392–394. doi: 10.1007/s00395-004-0483-6. [DOI] [PubMed] [Google Scholar]
- 109.Eaton P, Awad WI, Miller JI, Hearse DJ, Shattock MJ. Ischemic preconditioning: a potential role for constitutive low molecular weight stress protein translocation and phosphorylation? Journal of Molecular and Cellular Cardiology. 2000;32(6):961–971. doi: 10.1006/jmcc.2000.1136. [DOI] [PubMed] [Google Scholar]
- 110.Lewis SE, Mannion RJ, White FA, et al. A role for HSP27 in sensory neuron survival. Journal of Neuroscience. 1999;19(20):8945–8953. doi: 10.1523/JNEUROSCI.19-20-08945.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Brundel BJJM, Ke L, Dijkhuis AJ, et al. Heat shock proteins as molecular _targets for intervention in atrial fibrillation. Cardiovascular Research. 2008;78(3):422–428. doi: 10.1093/cvr/cvn060. [DOI] [PubMed] [Google Scholar]
- 112.Ke L, Meijering RAM, Hoogstra-Berends F, et al. HSPB1, HSPB6, HSPB7 and HSPB8 Protect against RhoA GTPase-Induced Remodeling in Tachypaced Atrial Myocytes. PLoS One. 2011;6(6) doi: 10.1371/journal.pone.0020395.e20395 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Wyttenbach A, Sauvageot O, Carmichael J, Diaz-Latoud C, Arrigo AP, Rubinsztein DC. Heat shock protein 27 prevents cellular polyglutamine toxicity and suppresses the increase of reactive oxygen species caused by huntingtin. Human Molecular Genetics. 2002;11(9):1137–1151. doi: 10.1093/hmg/11.9.1137. [DOI] [PubMed] [Google Scholar]
- 114.Outeiro TF, Klucken J, Strathearn KE, et al. Small heat shock proteins protect against α-synuclein-induced toxicity and aggregation. Biochemical and Biophysical Research Communications. 2006;351(3):631–638. doi: 10.1016/j.bbrc.2006.10.085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Lee S, Carson K, Rice-Ficht A, Good T. Small heat shock proteins differentially affect Aβ aggregation and toxicity. Biochemical and Biophysical Research Communications. 2006;347(2):527–533. doi: 10.1016/j.bbrc.2006.06.128. [DOI] [PubMed] [Google Scholar]
- 116.Perrin V, Régulier E, Abbas-Terki T, et al. Neuroprotection by Hsp104 and Hsp27 in lentiviral-based rat models of Huntington’s disease. Molecular Therapy. 2007;15(5):903–911. doi: 10.1038/mt.sj.6300141. [DOI] [PubMed] [Google Scholar]
- 117.Wilhelmus MMM, Boelens WC, Otte-Höller I, et al. Small heat shock protein HspB8: Its distribution in Alzheimer’s disease brains and its inhibition of amyloid-β protein aggregation and cerebrovascular amyloid-β toxicity. Acta Neuropathologica. 2006;111(2):139–149. doi: 10.1007/s00401-005-0030-z. [DOI] [PubMed] [Google Scholar]
- 118.Andley UP. Crystallins in the eye: function and pathology. Progress in Retinal and Eye Research. 2007;26(1):78–98. doi: 10.1016/j.preteyeres.2006.10.003. [DOI] [PubMed] [Google Scholar]
- 119.Vicart P, Caron A, Guicheney P, et al. A missense mutation in the αb-crystallin chaperone gene causes a desmin-related myopathy. Nature Genetics. 1998;20(1):92–95. doi: 10.1038/1765. [DOI] [PubMed] [Google Scholar]
- 120.Bova MP, Yaron O, Huang Q, et al. Mutation R120G in αB-crystallin, which is linked to a desmin-related myopathy, results in an irregular structure and defective chaperone-like function. Proceedings of the National Academy of Sciences of the United States of America. 1999;96(11):6137–6142. doi: 10.1073/pnas.96.11.6137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Evgrafov OV, Mersiyanova I, Irobi J, et al. Mutant small heat-shock protein 27 causes axonal Charcot-Marie-Tooth disease and distal hereditary motor neuropathy. Nature Genetics. 2004;36(6):602–606. doi: 10.1038/ng1354. [DOI] [PubMed] [Google Scholar]
- 122.Ackerley S, James PA, Kalli A, French S, Davies KE, Talbot K. A mutation in the small heat-shock protein HSPB1 leading to distal hereditary motor neuronopathy disrupts neurofilament assembly and the axonal transport of specific cellular cargoes. Human Molecular Genetics. 2006;15(2):347–354. doi: 10.1093/hmg/ddi452. [DOI] [PubMed] [Google Scholar]
- 123.Kijima K, Numakura C, Goto T, et al. Small heat shock protein 27 mutation in a Japanese patient with distal hereditary motor neuropathy. Journal of Human Genetics. 2005;50(9):473–476. doi: 10.1007/s10038-005-0280-6. [DOI] [PubMed] [Google Scholar]
- 124.Elicker KS, Hutson LD. Genome-wide analysis and expression profiling of the small heat shock proteins in zebrafish. Gene. 2007;403(1-2):60–69. doi: 10.1016/j.gene.2007.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Dierick I, Irobi J, Janssens S, et al. Genetic variant in the HSPB1 promoter region impairs the HSP27 stress response. Human Mutation. 2007;28(8):p. 830. doi: 10.1002/humu.9503. [DOI] [PubMed] [Google Scholar]
- 126.Ojha J, Masilamoni G, Dunlap D, Udoff RA, Cashikar AG. Sequestration of toxic oligomers by HspB1 as a cytoprotective mechanism. Molecular and Cellular Biology. 2011;31(15):3146–3157. doi: 10.1128/MCB.01187-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Vos MJ, Zijlstra MP, Kanon B, et al. HSPB7 is the most potent polyQ aggregation suppressor within the HSPB family of molecular chaperones. Human Molecular Genetics. 2010;19(23):4677–4693. doi: 10.1093/hmg/ddq398. [DOI] [PubMed] [Google Scholar]
- 128.Arrigo AP. Small stress proteins: chaperones that act as regulators of intracellular redox state and programmed cell death. Biological Chemistry. 1998;379(1):19–26. [PubMed] [Google Scholar]
- 129.Chen H, Zheng C, Zhang Y, Chang YZ, Qian ZM, Shen X. Heat shock protein 27 downregulates the transferrin receptor 1-mediated iron uptake. International Journal of Biochemistry and Cell Biology. 2006;38(8):1402–1416. doi: 10.1016/j.biocel.2006.02.006. [DOI] [PubMed] [Google Scholar]
- 130.Firdaus WJJ, Wyttenbach A, Giuliano P, Kretz-Remy C, Currie RW, Arrigo AP. Huntingtin inclusion bodies are iron-dependent centers of oxidative events. FEBS Journal. 2006;273(23):5428–5441. doi: 10.1111/j.1742-4658.2006.05537.x. [DOI] [PubMed] [Google Scholar]
- 131.Halliwell B. Role of free radicals in the neurodegenerative diseases: therapeutic implications for antioxidant treatment. Drugs and Aging. 2001;18(9):685–716. doi: 10.2165/00002512-200118090-00004. [DOI] [PubMed] [Google Scholar]
- 132.Bharath S, Hsu M, Kaur D, Rajagopalan S, Andersen JK. Glutathione, iron and Parkinson’s disease. Biochemical Pharmacology. 2002;64(5-6):1037–1048. doi: 10.1016/s0006-2952(02)01174-7. [DOI] [PubMed] [Google Scholar]
- 133.Jenner P, Olanow CW. Oxidative stress and the pathogenesis of Parkinson’s disease. Neurology. 1996;47(6):S161–S170. doi: 10.1212/wnl.47.6_suppl_3.161s. [DOI] [PubMed] [Google Scholar]
- 134.Browne SE, Ferrante RJ, Beal MF. Oxidative stress in Huntington’s disease. Brain Pathology. 1999;9(1):147–163. doi: 10.1111/j.1750-3639.1999.tb00216.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Tabner BJ, Turnbull S, El-Agnaf O, Allsop D. Production of reactive oxygen species from aggregating proteins implicated in Alzheimer’s disease, Parkinson’s disease and other neurodegenerative diseases. Current Topics in Medicinal Chemistry. 2001;1(6):507–517. doi: 10.2174/1568026013394822. [DOI] [PubMed] [Google Scholar]
- 136.Turnbull S, Tabner BJ, Brown DR, Allsop D. Copper-dependent generation of hydrogen peroxide from the toxic prion protein fragment PrP106-126. Neuroscience Letters. 2003;336(3):159–162. doi: 10.1016/s0304-3940(02)01254-5. [DOI] [PubMed] [Google Scholar]
- 137.Choi J, Rees HD, Weintraub ST, Levey AI, Chin LS, Li L. Oxidative modifications and aggregation of Cu,Zn-superoxide dismutase associated with alzheimer and Parkinson diseases. Journal of Biological Chemistry. 2005;280(12):11648–11655. doi: 10.1074/jbc.M414327200. [DOI] [PubMed] [Google Scholar]
- 138.Fox JH, Kama JA, Lieberman G, et al. Mechanisms of copper ion mediated Huntington’s disease progression. PLoS One. 2007;2(3, article e334) doi: 10.1371/journal.pone.0000334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Hilditch-Maguire P, Trettel F, Passani LA, Auerbach A, Persichetti F, MacDonald ME. Huntingtin: an iron-regulated protein essential for normal nuclear and perinuclear organelles. Human Molecular Genetics. 2000;9(19):2789–2797. doi: 10.1093/hmg/9.19.2789. [DOI] [PubMed] [Google Scholar]
- 140.Huang X, Moir RD, Tanzi RE, Bush AI, Rogers JT. Redox-active metals, oxidative stress, and Alzheimer’s disease pathology. Annals of the New York Academy of Sciences. 2004;1012:153–163. doi: 10.1196/annals.1306.012. [DOI] [PubMed] [Google Scholar]
- 141.Halliwell B, Gutteridge JMC. Role of iron in oxygen radical reactions. Methods in Enzymology. 1984;105:47–56. doi: 10.1016/s0076-6879(84)05007-2. [DOI] [PubMed] [Google Scholar]
- 142.Shoham S, Youdim MB. Iron involvement in neural damage and microgliosis in models of neurodegenerative diseases. Cellular and Molecular Biology. 2000;46(4):743–760. [PubMed] [Google Scholar]
- 143.Janué A, Olivé M, Ferrer I. Oxidative stress in desminopathies and myotilinopathies: a link between oxidative damage and abnormal protein aggregation. Brain Pathology. 2007;17(4):377–388. doi: 10.1111/j.1750-3639.2007.00087.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Liu J, Chen Q, Huang W, et al. Impairment of the ubiquitin-proteasome system in desminopathy mouse hearts. The FASEB Journal. 2006;20(2):362–364. doi: 10.1096/fj.05-4869fje. [DOI] [PubMed] [Google Scholar]
- 145.Mehlen P, Preville X, Chareyron P, Briolay J, Klemenz R, Arrigo AP. Constitutive expression of human hsp27, Drosophila hsp27, or human αB- crystallin confers resistance to TNF- and oxidative stress-induced cytotoxicity in stably transfected murine L929 fibroblasts. Journal of Immunology. 1995;154(1):363–374. [PubMed] [Google Scholar]
- 146.Kammanadiminti SJ, Chadee K. Suppression of NF-κB activation by Entamoeba histolytica in intestinal epithelial cells is mediated by heat shock protein 27. Journal of Biological Chemistry. 2006;281(36):26112–26120. doi: 10.1074/jbc.M601988200. [DOI] [PubMed] [Google Scholar]
- 147.Alford KA, Glennie S, Turrell BR, Rawlinson L, Saklatvala J, Dean JLE. Heat shock protein 27 functions in inflammatory gene expression and transforming growth factor-β-activated kinase-1 (TAK1)-mediated signaling. Journal of Biological Chemistry. 2007;282(9):6232–6241. doi: 10.1074/jbc.M610987200. [DOI] [PubMed] [Google Scholar]
- 148.Dodd SL, Hain B, Senf SM, Judge AR. Hsp27 inhibits IKKβ-induced NF-κB activity and skeletal muscle atrophy. The FASEB Journal. 2009;23(10):3415–3423. doi: 10.1096/fj.08-124602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Merendino AM, Paul C, Vignola AM, et al. Heat shock protein-27 protects human bronchial epithelial cells against oxidative stress-mediated apoptosis: possible implication in asthma. Cell Stress and Chaperones. 2002;7(3):269–280. doi: 10.1379/1466-1268(2002)007<0269:hspphb>2.0.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Dillmann WH. Heat shock proteins and protection against ischemic injury. Infectious Diseases in Obstetrics and Gynecology. 1999;7(1-2):55–57. doi: 10.1155/S1064744999000113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Charette SJ, Lavoie JN, Lambert H, Landry J. Inhibition of Daxx-mediated apoptosis by heat shock protein 27. Molecular and Cellular Biology. 2000;20(20):7602–7612. doi: 10.1128/mcb.20.20.7602-7612.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Pandey P, Farber R, Nakazawa A, et al. Hsp27 functions as a negative regulator of cytochrome c-dependent activation of procaspase-3. Oncogene. 2000;19(16):1975–1981. doi: 10.1038/sj.onc.1203531. [DOI] [PubMed] [Google Scholar]
- 153.Rane MJ, Pan Y, Singh S, et al. Heat shock protein 27 controls apoptosis by regulating Akt activation. Journal of Biological Chemistry. 2003;278(30):27828–27835. doi: 10.1074/jbc.M303417200. [DOI] [PubMed] [Google Scholar]
- 154.Kamradt MC, Lu M, Werner ME, et al. The small heat shock protein αB-crystallin is a novel inhibitor of TRAIL-induced apoptosis that suppresses the activation of caspase-3. Journal of Biological Chemistry. 2005;280(12):11059–11066. doi: 10.1074/jbc.M413382200. [DOI] [PubMed] [Google Scholar]
- 155.Wu R, Kausar H, Johnson P, Montoya-Durango DE, Merchant M, Rane MJ. Hsp27 regulates Akt activation and polymorphonuclear leukocyte apoptosis by scaffolding MK2 to Akt signal complex. Journal of Biological Chemistry. 2007;282(30):21598–21608. doi: 10.1074/jbc.M611316200. [DOI] [PubMed] [Google Scholar]
- 156.Garrido C, Bruey JM, Fromentin A, Hammann A, Arrigo AP, Solary E. HSP27 inhibits cytochrome C-dependent activation of procaspase-9. The FASEB Journal. 1999;13(14):2061–2070. doi: 10.1096/fasebj.13.14.2061. [DOI] [PubMed] [Google Scholar]
- 157.Bruey JM, Ducasse C, Bonniaud P, et al. Hsp27 negatively regulates cell death by interacting with cytochrome c. Nature Cell Biology. 2000;2(9):645–652. doi: 10.1038/35023595. [DOI] [PubMed] [Google Scholar]
- 158.Samali A, Robertson JD, Peterson E, et al. Hsp27 protects mitochondria of thermotolerant cells against apoptotic stimuli. Cell Stress and Chaperones. 2001;6(1):49–58. doi: 10.1379/1466-1268(2001)006<0049:hpmotc>2.0.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Paul C, Manero F, Gonin S, Kretz-Remy C, Virot S, Arrigo AP. Hsp27 as a negative regulator of cytochrome c release. Molecular and Cellular Biology. 2002;22(3):816–834. doi: 10.1128/MCB.22.3.816-834.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Mao YW, Liu JP, Xiang H, Li DWC. Human αA- and αB-crystallins bind to Bax and Bcl-Xs to sequester their translocation during staurosporine-induced apoptosis. Cell Death and Differentiation. 2004;11(5):512–526. doi: 10.1038/sj.cdd.4401384. [DOI] [PubMed] [Google Scholar]
- 161.Li DWC, Liu JP, Mao YW, et al. Calcium-activated RAF/MEK/ERK signaling pathway mediates p53-dependent apoptosis and is abrogated by αB-crystallin through inhibition of RAS activation. Molecular Biology of the Cell. 2005;16(9):4437–4453. doi: 10.1091/mbc.E05-01-0010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Havasi A, Li Z, Wang Z, et al. Hsp27 inhibits Bax activation and apoptosis via a phosphatidylinositol 3-kinase-dependent mechanism. Journal of Biological Chemistry. 2008;283(18):12305–12313. doi: 10.1074/jbc.M801291200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Arrigo A-P. Structure-functions of HspB1 (Hsp27) Methods in Molecular Biology. 2011;787:105–119. doi: 10.1007/978-1-61779-295-3_9. [DOI] [PubMed] [Google Scholar]
- 164.Rocchi P, Beraldi E, Ettinger S, et al. Increased Hsp27 after androgen ablation facilitates androgen-independent progression in prostate cancer via signal transducers and activators of transcription 3-mediated suppression of apoptosis. Cancer Research. 2005;65(23):11083–11093. doi: 10.1158/0008-5472.CAN-05-1840. [DOI] [PubMed] [Google Scholar]
- 165.Andrieu C, Taieb D, Baylot V, et al. Heat shock protein 27 confers resistance to androgen ablation and chemotherapy in prostate cancer cells through eIF4E. Oncogene. 2010;29(13):1883–1896. doi: 10.1038/onc.2009.479. [DOI] [PubMed] [Google Scholar]
- 166.Cayado-Gutiérrez N, Moncalero VL, Rosales EM, et al. Down-regulation of Hsp27 (HSPB1) in MCF-7 human breast cancer cells induces up-regulation of PTEN . doi: 10.1007/s12192-012-0367-x. Cell Stress and Chaperones. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Rane MJ, Coxon PY, Powell DW, et al. p38 Kinase-dependent MAPKAPK-2 Activation Functions as 3-Phosphoinositide-dependent Kinase-2 for Akt in Human Neutrophils. Journal of Biological Chemistry. 2001;276(5):3517–3523. doi: 10.1074/jbc.M005953200. [DOI] [PubMed] [Google Scholar]
- 168.Jomary C, Cullen J, Jones SE. Inactivation of the akt survival pathway during photoreceptor apoptosis in the retinal degeneration mouse. Investigative Ophthalmology and Visual Science. 2006;47(4):1620–1629. doi: 10.1167/iovs.05-1176. [DOI] [PubMed] [Google Scholar]
- 169.Hayashi N, Peacock JW, Beraldi E, Zoubeidi A, Gleave ME, Ong CJ. Hsp27 silencing coordinately inhibits proliferation and promotes Fas-induced apoptosis by regulating the PEA-15 molecular switch. Cell Death and Differentiation. 2012;19(6):990–1002. doi: 10.1038/cdd.2011.184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Kamradt MC, Chen F, Cryns VL. The small heat shock protein αB-crystallin negatively regulates cytochrome c- and caspase-8-dependent activation of caspase-3 by inhibiting its autoproteolytic maturation. Journal of Biological Chemistry. 2001;276(19):16059–16063. doi: 10.1074/jbc.C100107200. [DOI] [PubMed] [Google Scholar]
- 171.Kamradt MC, Chen F, Sam S, Cryns VL. The small heat shock protein αB-crystallin negatively regulates apoptosis during myogenic differentiation by inhibiting caspase-3 activation. Journal of Biological Chemistry. 2002;277(41):38731–38736. doi: 10.1074/jbc.M201770200. [DOI] [PubMed] [Google Scholar]
- 172.Liu JP, Schlosser R, Ma WY, et al. Human αA- and αB-crystallins prevent UVA-induced apoptosis through regulation of PKCα, RAF/MEK/ERK and AKT signaling pathways. Experimental Eye Research. 2004;79(3):393–403. doi: 10.1016/j.exer.2004.06.015. [DOI] [PubMed] [Google Scholar]
- 173.Launay N, Tarze A, Vicart P, Lilienbaum A. Serine 59 phosphorylation of αB-crystallin down-regulates its anti-apoptotic function by binding and sequestering Bcl-2 in breast cancer cells. Journal of Biological Chemistry. 2010;285(48):37324–37332. doi: 10.1074/jbc.M110.124388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Ciocca DR, Calderwood SK. Heat shock proteins in cancer: diagnostic, prognostic, predictive, and treatment implications. Cell Stress and Chaperones. 2005;10(2):86–103. doi: 10.1379/CSC-99r.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Calderwood SK, Khaleque MA, Sawyer DB, Ciocca DR. Heat shock proteins in cancer: chaperones of tumorigenesis. Trends in Biochemical Sciences. 2006;31(3):164–172. doi: 10.1016/j.tibs.2006.01.006. [DOI] [PubMed] [Google Scholar]
- 176.Arrigo AP, Gibert B. HspB1 dynamic phospho-oligomeric structure dependent interactome as cancer therapeutic _target. Current Molecular Medicine. 2012;12(9):1151–1163. doi: 10.2174/156652412803306693. [DOI] [PubMed] [Google Scholar]
- 177.Ciocca DR, Arrigo AP, Calderwood SK. Heat shock proteins and heat shock factor 1 in carcinogenesis and tumor development: an update. doi: 10.1007/s00204-012-0918-z. Archives of Toxicology. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Calderwood SK. HSF1, a versatile factor in tumorogenesis. Current Molecular Medicine. 2012;12(9):1102–1107. doi: 10.2174/156652412803306675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Santagata S, Hu R, Lin NU, et al. High levels of nuclear heat-shock factor 1 (HSF1) are associated with poor prognosis in breast cancer. Proceedings of the National Academy of Sciences of the United States of America. 2011;108(45):18378–18383. doi: 10.1073/pnas.1115031108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Trepel J, Mollapour M, Giaccone G, Neckers L. _targeting the dynamic HSP90 complex in cancer. Nature Reviews Cancer. 2010;10(8):537–549. doi: 10.1038/nrc2887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Mehlen P, Schulze-Osthoff K, Arrigo AP. Small stress proteins as novel regulators of apoptosis. Heat shock protein 27 blocks Fas/APO-1- and staurosporine-induced cell death. Journal of Biological Chemistry. 1996;271(28):16510–16514. doi: 10.1074/jbc.271.28.16510. [DOI] [PubMed] [Google Scholar]
- 182.Garrido C, Brunet M, Didelot C, Zermati Y, Schmitt E, Kroemer G. Heat shock proteins 27 and 70: anti-apoptotic proteins with tumorigenic properties. Cell Cycle. 2006;5(22):2592–2601. doi: 10.4161/cc.5.22.3448. [DOI] [PubMed] [Google Scholar]
- 183.O’Callaghan-Sunol C, Gabai VL, Sherman MY. Hsp27 modulates p53 signaling and suppresses cellular senescence. Cancer Research. 2007;67(24):11779–11788. doi: 10.1158/0008-5472.CAN-07-2441. [DOI] [PubMed] [Google Scholar]
- 184.Zhuang H, Jiang W, Cheng W, et al. Down-regulation of HSP27 sensitizes TRAIL-resistant tumor cell to TRAIL-induced apoptosis. Lung Cancer. 2010;68(1):27–38. doi: 10.1016/j.lungcan.2009.05.014. [DOI] [PubMed] [Google Scholar]
- 185.Garrido C, Fromentin A, Bonnotte B, et al. Heat shock protein 27 enhances the tumorigenicity of immunogenic rat colon carcinoma cell clones. Cancer Research. 1998;58(23):5495–5499. [PubMed] [Google Scholar]
- 186.Bruey JM, Paul C, Fromentin A, et al. Differential regulation of HSP27 oligomerization in tumor cells grown in vitro and in vivo. Oncogene. 2000;19(42):4855–4863. doi: 10.1038/sj.onc.1203850. [DOI] [PubMed] [Google Scholar]
- 187.Lemieux P, Oesterreich S, Lawrence JA, et al. The small heat shock protein hsp27 increases invasiveness but decreases motility of breast cancer cells. Invasion and Metastasis. 1997;17(3):113–123. [PubMed] [Google Scholar]
- 188.Bausero MA, Bharti A, Page DT, et al. Silencing the hsp25 gene eliminates migration capability of the highly metastatic murine 4T1 breast adenocarcinoma cell. Tumor Biology. 2006;27(1):17–26. doi: 10.1159/000090152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Gibert B, Eckel B, Gonin V, et al. _targeting heat shock protein 27 (HspB1) interferes with bone metastasis and tumour formation in vivo. British Journal of Cancer. 2012;107(1):63–70. doi: 10.1038/bjc.2012.188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Nagaraja GN, Kaur P, Asea A. Role of human and mouse HspB1 in metastasis. Current Molecular Medicine. 2012;12(9):1142–1150. doi: 10.2174/156652412803306701. [DOI] [PubMed] [Google Scholar]
- 191.Xu L, Chen S, Bergan RC. MAPKAPK2 and HSP27 are downstream effectors of p38 MAP kinase-mediated matrix metalloproteinase type 2 activation and cell invasion in human prostate cancer. Oncogene. 2006;25(21):2987–2998. doi: 10.1038/sj.onc.1209337. [DOI] [PubMed] [Google Scholar]
- 192.Fanelli MA, Montt-Guevara M, Diblasi AM, et al. P-Cadherin and β-catenin are useful prognostic markers in breast cancer patients; β-catenin interacts with heat shock protein Hsp27. Cell Stress and Chaperones. 2008;13(2):207–220. doi: 10.1007/s12192-007-0007-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Wei L, Liu TT, Wang HH, ", et al. Hsp27 participates in the maintenance of breast cancer stem cells through regulation of epithelial-mesenchymal transition and nuclear factor-kappaB, " Breast Cancer Research. 2011;13(5):p. R101. doi: 10.1186/bcr3042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Huot J, Roy G, Lambert H, Chretien P, Landry J. Increased survival after treatments with anticancer agents of Chinese hamster cells expressing the human M(r) 27,000 heat shock protein. Cancer Research. 1991;51(19):5245–5252. [PubMed] [Google Scholar]
- 195.Richards EH, Hickey E, Weber L, Masters JRW. Effect of overexpression of the small heat shock protein HSP27 on the heat and drug sensitivities of human testis tumor cells. Cancer Research. 1996;56(10):2446–2451. [PubMed] [Google Scholar]
- 196.Kamada M, So A, Muramaki M, Rocchi P, Beraldi E, Gleave M. Hsp27 knockdown using nucleotide-based therapies inhibit tumor growth and enhance chemotheraphy in human bladder cancer cells. Molecular Cancer Therapeutics. 2007;6(1):299–308. doi: 10.1158/1535-7163.MCT-06-0417. [DOI] [PubMed] [Google Scholar]
- 197.Kang SH, Kang KW, Kim KH, et al. Upregulated HSP27 in human breast cancer cells reduces Herceptin susceptibility by increasing Her2 protein stability. BMC Cancer. 2008;8, article 286 doi: 10.1186/1471-2407-8-286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Kase S, Parikh JG, Rao NA. Expression of heat shock protein 27 and alpha-crystallins in human retinoblastoma after chemoreduction. British Journal of Ophthalmology. 2009;93(4):541–544. doi: 10.1136/bjo.2008.145508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Gruvberger-Saal SK, Parsons R. Is the small heat shock protein αB-crystallin an oncogene? Journal of Clinical Investigation. 2006;116(1):30–32. doi: 10.1172/JCI27462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Moyano JV, Evans JR, Chen F, et al. αB-Crystallin is a novel oncoprotein that predicts poor clinical outcome in breast cancer. Journal of Clinical Investigation. 2006;116(1):261–270. doi: 10.1172/JCI25888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Deng M, Chen PC, Xie S, et al. The small heat shock protein α A-crystallin is expressed in pancreas and acts as a negative regulator of carcinogenesis. Biochimica et Biophysica Acta. 2010;1802(7-8):621–631. doi: 10.1016/j.bbadis.2010.04.004. [DOI] [PubMed] [Google Scholar]
- 202.Pockley AG, Multhoff G. Cell stress proteins in extracellular fluids: friend or foe? Novartis Foundation Symposium. 2008;291:86–95. doi: 10.1002/9780470754030.ch7. [DOI] [PubMed] [Google Scholar]
- 203.de Maio A. Extracellular heat shock proteins, cellular export vesicles, and the stress observation system: a form of communication during injury, infection, and cell damage: it is never known how far a controversial finding will go! Dedicated to Ferruccio Ritossa. Cell Stress and Chaperones. 2011;16(3):235–249. doi: 10.1007/s12192-010-0236-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Tytell M, Greenberg SG, Lasek RJ. Heat shock-like protein is transferred from glia to axon. Brain Research. 1986;363(1):161–164. doi: 10.1016/0006-8993(86)90671-2. [DOI] [PubMed] [Google Scholar]
- 205.Xu Q, Schett G, Seitz CS, Hu Y, Gupta RS, Wick G. Surface staining and cytotoxic activity of heat-shock protein 60 antibody in stressed aortic endothelial cells. Circulation Research. 1994;75(6):1078–1085. doi: 10.1161/01.res.75.6.1078. [DOI] [PubMed] [Google Scholar]
- 206.Multhoff G, Hightower LE. Cell surface expression of heat shock proteins and the immune response. Cell Stress and Chaperones. 1996;1(3):167–176. doi: 10.1379/1466-1268(1996)001<0167:cseohs>2.3.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Multhoff G. Heat shock protein 70 (Hsp70): membrane location, export and immunological relevance. Methods. 2007;43(3):229–237. doi: 10.1016/j.ymeth.2007.06.006. [DOI] [PubMed] [Google Scholar]
- 208.Bausero MA, Gastpar R, Multhoff G, Asea A. Alternative mechanism by which IFN-γ enhances tumor recognition: active release of heat shock protein 72. Journal of Immunology. 2005;175(5):2900–2912. doi: 10.4049/jimmunol.175.5.2900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Guzhova I, Kislyakova K, Moskaliova O, et al. In vitro studies show that Hsp70 can be released by glia and that exogenous Hsp70 can enhance neuronal stress tolerance. Brain Research. 2001;914(1-2):66–73. doi: 10.1016/s0006-8993(01)02774-3. [DOI] [PubMed] [Google Scholar]
- 210.Hightower LE, Guidon PT. Selective release from cultured mammalian cells of heat-shock (stress) proteins that resemble glia-axon transfer proteins. Journal of Cellular Physiology. 1989;138(2):257–266. doi: 10.1002/jcp.1041380206. [DOI] [PubMed] [Google Scholar]
- 211.Hunter-Lavin C, Davies EL, Bacelar MMFVG, Marshall MJ, Andrew SM, Williams JHH. Hsp70 release from peripheral blood mononuclear cells. Biochemical and Biophysical Research Communications. 2004;324(2):511–517. doi: 10.1016/j.bbrc.2004.09.075. [DOI] [PubMed] [Google Scholar]
- 212.Tytell M. Release of heat shock proteins (Hsps) and the effects of extracellular Hsps on neural cells and tissues. International Journal of Hyperthermia. 2005;21(5):445–455. doi: 10.1080/02656730500041921. [DOI] [PubMed] [Google Scholar]
- 213.Xu Q, Willeit J, Marosi M, et al. Association of serum antibodies to heat-shock protein 65 with carotid atherosclerosis. The Lancet. 1993;341(8840):255–259. doi: 10.1016/0140-6736(93)92613-x. [DOI] [PubMed] [Google Scholar]
- 214.Pockley AG, Shepherd J, Corton JM. Detection of heat shock protein 70 (Hsp70) and anti-Hsp70 antibodies in the serum of normal individuals. Immunological Investigations. 1998;27(6):367–377. doi: 10.3109/08820139809022710. [DOI] [PubMed] [Google Scholar]
- 215.Pockley AG, Bulmer J, Hanks BM, Wright BH. Identification of human heat shock protein 60 (Hsp60) and anti-Hsp60 antibodies in the peripheral circulation of normal individuals. Cell Stress and Chaperones. 1999;4(1):29–35. doi: 10.1054/csac.1998.0121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Walsh RC, Koukoulas I, Garnham A, Moseley PL, Hargreaves M, Febbraio MA. Exercise increases serum Hsp72 in humans. Cell Stress and Chaperones. 2001;6(4):386–393. doi: 10.1379/1466-1268(2001)006<0386:eishih>2.0.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Lewthwaite J, Owen N, Coates A, Henderson B, Steptoe A. Circulating human heat shock protein 60 in the plasma of British civil servants: relationship to physiological and psychosocial stress. Circulation. 2002;106(2):196–201. doi: 10.1161/01.cir.0000021121.26290.2c. [DOI] [PubMed] [Google Scholar]
- 218.Wright BH, Corton JM, El-Nahas AM, Wood RFM, Pockley AG. Elevated levels of circulating heat shock protein 70 (Hsp70) in peripheral and renal vascular disease. Heart and Vessels. 2000;15(1):18–22. doi: 10.1007/s003800070043. [DOI] [PubMed] [Google Scholar]
- 219.Campisi J, Leem TH, Fleshner M. Stress-induced extracellular Hsp72 is a functionally significant danger signal to the immune system. Cell Stress and Chaperones. 2003;8(3):272–286. doi: 10.1379/1466-1268(2003)008<0272:sehiaf>2.0.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Novoselova TV, Margulis BA, Novoselov SS, et al. Treatment with extracellular HSP70/HSC70 protein can reduce polyglutamine toxicity and aggregation. Journal of Neurochemistry. 2005;94(3):597–606. doi: 10.1111/j.1471-4159.2005.03119.x. [DOI] [PubMed] [Google Scholar]
- 221.Andreeva LI, Shabanov PD, Margulis BA. Exogenous heat shock protein with a molecular weight of 70 kDa changes behavior in white rats. Doklady Biological Sciences. 2004;394:34–37. doi: 10.1023/b:dobs.0000017124.29291.d5. [DOI] [PubMed] [Google Scholar]
- 222.Pockley AG, Georgiades A, Thulin T, de Faire U, Frostegård J. Serum heat shock protein 70 levels predict the development of atherosclerosis in subjects with established hypertension. Hypertension. 2003;42(3):235–238. doi: 10.1161/01.HYP.0000086522.13672.23. [DOI] [PubMed] [Google Scholar]
- 223.Kocsis J, Madaras B, Tóth EK, Füst G, Prohászka Z. Serum level of soluble 70-kD heat shock protein is associated with high mortality in patients with colorectal cancer without distant metastasis. Cell Stress and Chaperones. 2010;15(2):143–151. doi: 10.1007/s12192-009-0128-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.van Eden W, Koets A, van Kooten P, Prakken B, van der Zee R. Immunopotentiating heat shock proteins: negotiators between innate danger and control of autoimmunity. Vaccine. 2003;21(9-10):897–901. doi: 10.1016/s0264-410x(02)00538-8. [DOI] [PubMed] [Google Scholar]
- 225.Prohászka Z, Füst G. Immunological aspects of heat-shock proteins—the optimum stress of life. Molecular Immunology. 2004;41(1):29–44. doi: 10.1016/j.molimm.2004.02.001. [DOI] [PubMed] [Google Scholar]
- 226.Srivastava P. Roles of heat-shock proteins in innate and adaptive immunity. Nature Reviews Immunology. 2002;2(3):185–194. doi: 10.1038/nri749. [DOI] [PubMed] [Google Scholar]
- 227.Kurotaki T, Tamura Y, Ueda G, et al. Efficient cross-presentation by heat shock protein 90-peptide complex-loaded dendritic cells via an endosomal pathway. Journal of Immunology. 2007;179(3):1803–1813. doi: 10.4049/jimmunol.179.3.1803. [DOI] [PubMed] [Google Scholar]
- 228.Tamura Y, Torigoe T, Kutomi G, Hirata K, Sato N. New paradigm for intrinsic function of heat shock proteins as endogenous ligands in inflammation and innate immunity. Current Molecular Medicine. 2012;12(9):1198–1206. doi: 10.2174/156652412803306710. [DOI] [PubMed] [Google Scholar]
- 229.Ciocca DR, Frayssinet P, Cuello-Carrión FD. A pilot study with a therapeutic vaccine based on hydroxyapatite ceramic particles and self-antigens in cancer patients. Cell Stress and Chaperones. 2007;12(1):33–43. doi: 10.1379/CSC-218R.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Ciocca DR, Cayado-Guiterrez N, Maccioni M, Cuello-Carrio FD. Heat Shock Proteins (HSPs) based anti-cancer vaccines. Current Molecular Medicine. 2012;12(9):1183–1197. doi: 10.2174/156652412803306684. [DOI] [PubMed] [Google Scholar]
- 231.Binder RJ, Vatner R, Srivastava P. The heat-shock protein receptors: some answers and more questions. Tissue Antigens. 2004;64(4):442–451. doi: 10.1111/j.1399-0039.2004.00299.x. [DOI] [PubMed] [Google Scholar]
- 232.Ueda G, Tamura Y, Hirai I, et al. Tumor-derived heat shock protein 70-pulsed dendritic cells elicit-tumor-specific cytotoxic T lymphocytes (CTLs) and tumor immunity. Cancer Science. 2004;95(3):248–253. doi: 10.1111/j.1349-7006.2004.tb02211.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Mehlen P, Arrigo AP. The serum-induced phosphorylation of mammalian hsp27 correlates with changes in its intracellular localization and levels of oligomerization. European Journal of Biochemistry. 1994;221(1):327–334. doi: 10.1111/j.1432-1033.1994.tb18744.x. [DOI] [PubMed] [Google Scholar]
- 234.Tsvetkova NM, Horváth I, Török Z, et al. Small heat-shock proteins regulate membrane lipid polymorphism. Proceedings of the National Academy of Sciences of the United States of America. 2002;99(21):13504–13509. doi: 10.1073/pnas.192468399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Chowdary TK, Bakthisaran R, Tangirala R, Rao MC. Interaction of mammalian Hsp22 with lipid membranes. Biochemical Journal. 2007;401(2):437–445. doi: 10.1042/BJ20061046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Rayner K, Chen YX, McNulty M, et al. Extracellular release of the atheroprotective heat shock protein 27 is mediated by estrogen and competitively inhibits acLDL binding to scavenger receptor-a. Circulation Research. 2008;103(2):133–141. doi: 10.1161/CIRCRESAHA.108.172155. [DOI] [PubMed] [Google Scholar]
- 237.Rayner K, Sun J, Chen YX, et al. Heat shock protein 27 protects against atherogenesis via an estrogen-dependent mechanism: role of selective estrogen receptor beta modulation. Arteriosclerosis, Thrombosis, and Vascular Biology. 2009;29(11):1751–1756. doi: 10.1161/ATVBAHA.109.193656. [DOI] [PubMed] [Google Scholar]
- 238.Rayner K, Chen YX, Siebert T, O’Brien ER. Heat shock protein 27: clue to understanding estrogen-mediated atheroprotection? Trends in Cardiovascular Medicine. 2010;20(2):54–58. doi: 10.1016/j.tcm.2010.03.008. [DOI] [PubMed] [Google Scholar]
- 239.Salari S, Seibert T, Chen YX, et al. Extracellular HSP27 acts as a signaling molecule to activate NF-κB in macrophages. doi: 10.1007/s12192-012-0356-0. Cell Stress and Chaperones. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Henderson B, Graham Pockley A. Proteotoxic stress and circulating cell stress proteins in the cardiovascular diseases. Cell Stress and Chaperones. 2012;17(3):303–311. doi: 10.1007/s12192-011-0318-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Bausero MA, Page DT, Osinaga E, Asea A. Surface expression of Hsp25 and Hsp72 differentially regulates tumor growth and metastasis. Tumor Biology. 2004;25(5-6):243–251. doi: 10.1159/000081387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.Korneeva I, Caputo TA, Witkin SS. Cell-free 27 kDa heat shock protein (HSP27) and HSP27-cytochrome C complexes in the cervix of women with ovarian or endometrial cancer. International Journal of Cancer. 2002;102(5):483–486. doi: 10.1002/ijc.10747. [DOI] [PubMed] [Google Scholar]
- 243.Dempsey NC, Leoni F, Ireland HE, Hoyle C, Williams JHH. Differential heat shock protein localization in chronic lymphocytic leukemia. Journal of Leukocyte Biology. 2010;87(3):467–476. doi: 10.1189/jlb.0709502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244.Laudanski K, De A, Miller-Graziano C. Exogenous heat shock protein 27 uniquely blocks differentiation of monocytes to dendritic cells. European Journal of Immunology. 2007;37(10):2812–2824. doi: 10.1002/eji.200636993. [DOI] [PubMed] [Google Scholar]
- 245.Gruden G, Bruno G, Chaturvedi N, et al. Serum heat shock protein 27 and diabetes complications in the EURODIAB prospective complications study: a novel circulating marker for diabetic neuropathy. Diabetes. 2008;57(7):1966–1970. doi: 10.2337/db08-0009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246.Evdonin AL, Martynova MG, Bystrova OA, Guzhova IV, Margulis BA, Medvedeva ND. The release of Hsp70 from A431 carcinoma cells is mediated by secretory-like granules. European Journal of Cell Biology. 2006;85(6):443–455. doi: 10.1016/j.ejcb.2006.02.008. [DOI] [PubMed] [Google Scholar]
- 247.Vega VL, Rodríguez-Silva M, Frey T, et al. Hsp70 translocates into the plasma membrane after stress and is released into the extracellular environment in a membrane-associated form that activates macrophages. Journal of Immunology. 2008;180(6):4299–4307. doi: 10.4049/jimmunol.180.6.4299. [DOI] [PubMed] [Google Scholar]
- 248.Hurwitz MD, Kaur P, Nagaraja GM, Bausero MA, Manola J, Asea A. Radiation therapy induces circulating serum Hsp72 in patients with prostate cancer. Radiotherapy and Oncology. 2010;95(3):350–358. doi: 10.1016/j.radonc.2010.03.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Gastpar R, Gehrmann M, Bausero MA, et al. Heat shock protein 70 surface-positive tumor exosomes stimulate migratory and cytolytic activity of natural killer cells. Cancer Research. 2005;65(12):5238–5247. doi: 10.1158/0008-5472.CAN-04-3804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250.Liebhardt S, Ditsch N, Nieuwland R, et al. CEA-, Her2/neu-, BCRP- and Hsp27-positive microparticles in breast cancer patients. Anticancer Research. 2010;30(5):1707–1712. [PubMed] [Google Scholar]
- 251.Calderwood SK, Mambula SS, Gray PJ. Extracellular heat shock proteins in cell signaling and immunity. Annals of the New York Academy of Sciences. 2007;1113:28–39. doi: 10.1196/annals.1391.019. [DOI] [PubMed] [Google Scholar]
- 252.Asea A, Kabingu E, Stevenson MA, Calderwood SK. HSP70 peptidembearing and peptide-negative preparations act as chaperokines. Cell Stress and Chaperones. 2000;5(5):425–431. doi: 10.1379/1466-1268(2000)005<0425:hpbapn>2.0.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Asea A, Kraeft SK, Kurt-Jones EA, et al. HSP70 stimulates cytokine production through a CD 14-dependant pathway, demonstrating its dual role as a chaperone and cytokine. Nature Medicine. 2000;6(4):435–442. doi: 10.1038/74697. [DOI] [PubMed] [Google Scholar]
- 254.Asea A, Rehli M, Kabingu E, et al. Novel signal transduction pathway utilized by extracellular HSP70. Role of toll-like receptor (TLR) 2 and TLR4. Journal of Biological Chemistry. 2002;277(17):15028–15034. doi: 10.1074/jbc.M200497200. [DOI] [PubMed] [Google Scholar]
- 255.Lewthwaite J, George R, Lund PA, et al. Rhizobium leguminosarum chaperonin 60.3, but not chaperonin 60.1, induces cytokine production by human monocytes: activity is dependent on interaction with cell surface CD14. Cell Stress and Chaperones. 2002;7(2):130–136. doi: 10.1379/1466-1268(2002)007<0130:rlcbnc>2.0.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 256.Delneste Y, Magistrelli G, Gauchat JF, et al. Involvement of LOX-1 in dendritic cell-mediated antigen cross-presentation. Immunity. 2002;17(3):353–362. doi: 10.1016/s1074-7613(02)00388-6. [DOI] [PubMed] [Google Scholar]
- 257.Thériault JR, Adachi H, Calderwood SK. Role of scavenger receptors in the binding and internalization of heat shock protein 70. Journal of Immunology. 2006;177(12):8604–8611. doi: 10.4049/jimmunol.177.12.8604. [DOI] [PubMed] [Google Scholar]
- 258.Berwin B, Hart JP, Rice S, et al. Scavenger receptor-A mediates gp96/GRP94 and calreticulin internalization by antigen-presenting cells. The EMBO Journal. 2003;22(22):6127–6136. doi: 10.1093/emboj/cdg572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 259.Wang XY, Facciponte J, Chen X, Subjeck JR, Repasky EA. Scavenger receptor-A negatively regulates antitumor immunity. Cancer Research. 2007;67(10):4996–5002. doi: 10.1158/0008-5472.CAN-06-3138. [DOI] [PubMed] [Google Scholar]
- 260.Chalmin F, Ladoire S, Mignot G, et al. Membrane-associated Hsp72 from tumor-derived exosomes mediates STAT3-dependent immunosuppressive function of mouse and human myeloid-derived suppressor cells. Journal of Clinical Investigation. 2010;120(2):457–471. doi: 10.1172/JCI40483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 261.Wu T, Tanguay RM. Antibodies against heat shock proteins in environmental stresses and diseases: friend or foe? Cell Stress and Chaperones. 2006;11(1):1–12. doi: 10.1379/CSC-155R.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262.Conroy SE, Sasieni PD, Amin V, et al. Antibodies to heat-shock protein 27 are associated with improved survival in patients with breast cancer. British Journal of Cancer. 1998;77(11):1875–1879. doi: 10.1038/bjc.1998.312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263.Joachim SC, Bruns K, Lackner KJ, Pfeiffer N, Grus FH. Antibodies to α B-crystallin, vimentin, and heat shock protein 70 in aqueous humor of patients with normal tension glaucoma and IgG antibody patterns against retinal antigen in aqueous humor. Current Eye Research. 2007;32(6):501–509. doi: 10.1080/02713680701375183. [DOI] [PubMed] [Google Scholar]
- 264.Tezel G, Wax MB. Inhibition of caspase activity in retinal cell apoptosis induced by various stimuli in vitro. Investigative Ophthalmology and Visual Science. 1999;40(11):2660–2667. [PubMed] [Google Scholar]
- 265.Tezel G, Wax MB. The mechanisms of hsp27 antibody-mediated apoptosis in retinal neuronal cells. Journal of Neuroscience. 2000;20(10):3552–3562. doi: 10.1523/JNEUROSCI.20-10-03552.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 266.Besgen P, Trommler P, Vollmer S, Prinz JC. Ezrin, maspin, peroxiredoxin 2, and heat shock protein 27: potential _targets of a streptococcal-induced autoimmune response in psoriasis. Journal of Immunology. 2010;184(9):5392–5402. doi: 10.4049/jimmunol.0903520. [DOI] [PubMed] [Google Scholar]
- 267.Agius MA, Kirvan CA, Schafer AL, Gudipati E, Zhu S. High prevalence of anti-α-crystallin antibodies in multiple sclerosis: correlation with severity and activity of disease. Acta Neurologica Scandinavica. 1999;100(3):139–147. doi: 10.1111/j.1600-0404.1999.tb00729.x. [DOI] [PubMed] [Google Scholar]
- 268.Rothbard JB, Zhao X, Sharpe O, et al. Chaperone activity of α B-crystallin is responsible for its incorrect assignment as an autoantigen in multiple sclerosis. Journal of Immunology. 2011;186(7):4263–4268. doi: 10.4049/jimmunol.1003934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 269.Neckers L, Mimnaugh E, Schulte TW. Hsp90 as an anti-cancer _target. Drug Resistance Updates. 1999;2(3):165–172. doi: 10.1054/drup.1999.0082. [DOI] [PubMed] [Google Scholar]
- 270.Georgakis GV, Younes A. Heat-shock protein 90 inhibitors in cancer therapy: 17AAG and beyond. Future Oncology. 2005;1(2):273–281. doi: 10.1517/14796694.1.2.273. [DOI] [PubMed] [Google Scholar]
- 271.McClellan AJ, Xia Y, Deutschbauer AM, Davis RW, Gerstein M, Frydman J. Diverse cellular functions of the Hsp90 molecular chaperone uncovered using systems approaches. Cell. 2007;131(1):121–135. doi: 10.1016/j.cell.2007.07.036. [DOI] [PubMed] [Google Scholar]
- 272.Moulick K, Ahn JH, Zong H, et al. Affinity-based proteomics reveal cancer-specific networks coordinated by Hsp90. Nature Chemical Biology. 2011;7(11):818–826. doi: 10.1038/nchembio.670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 273.Hessling M, Richter K, Buchner J. Dissection of the ATP-induced conformational cycle of the molecular chaperone Hsp90. Nature Structural and Molecular Biology. 2009;16(3):287–293. doi: 10.1038/nsmb.1565. [DOI] [PubMed] [Google Scholar]
- 274.Mickler M, Hessling M, Ratzke C, Buchner J, Hugel T. The large conformational changes of Hsp90 are only weakly coupled to ATP hydrolysis. Nature Structural and Molecular Biology. 2009;16(3):281–286. doi: 10.1038/nsmb.1557. [DOI] [PubMed] [Google Scholar]
- 275.Gibert B, Hadchity E, Czekalla A, et al. Inhibition of heat shock protein 27 (HspB1) tumorigenic functions by peptide aptamers. Oncogene. 2011;30:3672–3681. doi: 10.1038/onc.2011.73. [DOI] [PubMed] [Google Scholar]
- 276.Stokoe D, Engel K, Campbell DG, Cohen P, Gaestel M. Identification of MAPKAP kinase 2 as a major enzyme responsible for the phosphorylation of the small mammalian heat shock proteins. FEBS Letters. 1992;313(3):307–313. doi: 10.1016/0014-5793(92)81216-9. [DOI] [PubMed] [Google Scholar]
- 277.Rouse J, Cohen P, Trigon S, et al. A novel kinase cascade triggered by stress and heat shock that stimulates MAPKAP kinase-2 and phosphorylation of the small heat shock proteins. Cell. 1994;78(6):1027–1037. doi: 10.1016/0092-8674(94)90277-1. [DOI] [PubMed] [Google Scholar]
- 278.Zantema A, Vries MV-D, Maasdam D, Bol S, Eb AVD. Heat shock protein 27 and αB-crystallin can form a complex, which dissociates by heat shock. Journal of Biological Chemistry. 1992;267(18):12936–12941. [PubMed] [Google Scholar]
- 279.Saha S, Das KP. Relationship between chaperone activity and oligomeric size of recombinant human αA- and αB-crystallin: a tryptic digestion study. Proteins. 2004;57(3):610–617. doi: 10.1002/prot.20230. [DOI] [PubMed] [Google Scholar]
- 280.Simon S, Fontaine JM, Martin JL, et al. Myopathy-associated αB-crystallin mutants: abnormal phosphorylation, intracellular location, and interactions with other small heat shock proteins. Journal of Biological Chemistry. 2007;282(47):34276–34287. doi: 10.1074/jbc.M703267200. [DOI] [PubMed] [Google Scholar]
- 281.Bukach OV, Glukhova AE, Seit-Nebi AS, Gusev NB. Heterooligomeric complexes formed by human small heat shock proteins HspB1 (Hsp27) and HspB6 (Hsp20) Biochimica et Biophysica Acta. 2009;1794(3):486–495. doi: 10.1016/j.bbapap.2008.11.010. [DOI] [PubMed] [Google Scholar]
- 282.Gidalevitz T, Ben-Zvi A, Ho KH, Brignull HR, Morimoto RI. Progressive disruption of cellular protein folding in models of polyglutamine diseases. Science. 2006;311(5766):1471–1474. doi: 10.1126/science.1124514. [DOI] [PubMed] [Google Scholar]
- 283.Sanbe A, Daicho T, Mizutani R, et al. Protective effect of geranylgeranylacetone via enhancement of HSPB8 induction in desmin-related cardiomyopathy. PLoS One. 2009;4(4) doi: 10.1371/journal.pone.0005351.e5351 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 284.Babizhayev MA. Current ocular drug delivery challenges for N-acetylcarnosine: novel patented routes and modes of delivery, design for enhancement of therapeutic activity and drug delivery relationships. Recent Patents on Drug Delivery and Formulation. 2009;3(3):229–265. doi: 10.2174/187221109789105621. [DOI] [PubMed] [Google Scholar]
- 285.Babizhayev MA, Burke L, Micans P, Richer SP. N-acetylcarnosine sustained drug delivery eye drops to control the signs of ageless vision: glare sensitivity, cataract amelioration and quality of vision currently available treatment for the challenging 50,000-patient population. Clinical Interventions in Aging. 2009;4(1):31–50. [PMC free article] [PubMed] [Google Scholar]
- 286.Gibert B, Simon S, Dimitrova V, Diaz-Latoud C, Arrigo AP. Peptide aptamers-tools to negatively or positively modulate HspB1(27) function. doi: 10.1098/rstb.2012.0075. Philosophical Transactions of The Royal Society B. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 287.Aloy MT, Hadchity E, Bionda C, et al. Protective role of Hsp27 protein against gamma radiation-induced apoptosis and radiosensitization effects of Hsp27 gene silencing in different human tumor cells. International Journal of Radiation Oncology Biology Physics. 2008;70(2):543–553. doi: 10.1016/j.ijrobp.2007.08.061. [DOI] [PubMed] [Google Scholar]
- 288.Rocchi P, Jugpal P, So A, et al. Small interference RNA _targeting heat-shock protein 27 inhibits the growth of prostatic cell lines and induces apoptosis via caspase-3 activation in vitro. BJU International. 2006;98(5):1082–1089. doi: 10.1111/j.1464-410X.2006.06425.x. [DOI] [PubMed] [Google Scholar]
- 289.Kaur P, Nagaraja GM, Asea A. Combined lentiviral and RNAi technologies for the delivery and permanent silencing of the hsp25 gene. Methods in Molecular Biology. 2011;787:121–136. doi: 10.1007/978-1-61779-295-3_10. [DOI] [PubMed] [Google Scholar]
- 290.Stengel F, Baldwin AJ, Painter AJ, et al. Quaternary dynamics and plasticity underlie small heat shock protein chaperone function. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(5):2007–2012. doi: 10.1073/pnas.0910126107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 291.Basha E, O'Neill H, Vierling E. Small heat shock proteins and α-crystallins: dynamic proteins with flexible functions. Trends in Biochemical Sciences. 2012;37(3):106–117. doi: 10.1016/j.tibs.2011.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 292.Heinrich JC, Tuukkanen A, Schroeder M, Fahrig T, Fahrig R. RP101 (brivudine) binds to heat shock protein HSP27 (HSPB1) and enhances survival in animals and pancreatic cancer patients. Journal of Cancer Research and Clinical Oncology. 2011;137(9):1349–1361. doi: 10.1007/s00432-011-1005-1. [DOI] [PubMed] [Google Scholar]
- 293.Bagnéris C, Bateman OA, Naylor CE, et al. Crystal structures of α-crystallin domain dimers of αB-crystallin and Hsp20. Journal of Molecular Biology. 2009;392(5):1242–1252. doi: 10.1016/j.jmb.2009.07.069. [DOI] [PubMed] [Google Scholar]
- 294.Bagnéris C, Bateman OA, Naylor CE, et al. Corrigendum to: Crystal structures of α-crystallin domain dimers of αB-crystallin and Hsp20. Journal of Molecular Biology. 2009;394(3):p. 588. doi: 10.1016/j.jmb.2009.07.069. [DOI] [PubMed] [Google Scholar]
- 295.Whitesell L, Santagata S, NU L. Inhibiting Hsp90 to treat cancer: a strategy in evolution. Current Molecular Medicine. 2012;12(9):1108–1124. doi: 10.2174/156652412803306657. [DOI] [PubMed] [Google Scholar]